
Neobisium ( Neobisium ) zoiai Gardini & Rizzerio, 1986
|
publication ID |
https://doi.org/10.11646/zootaxa.5381.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9B004CC6-C0AE-4205-A99C-A41AA14319E6 |
|
DOI |
https://doi.org/10.5281/zenodo.10392408 |
|
persistent identifier |
https://treatment.plazi.org/id/03883A10-B909-FFEE-FF54-BFD9FBDC222A |
|
treatment provided by |
Plazi |
|
scientific name |
Neobisium ( Neobisium ) zoiai Gardini & Rizzerio, 1986 |
| status |
|
Neobisium ( Neobisium) zoiai Gardini & Rizzerio, 1986 View in CoL
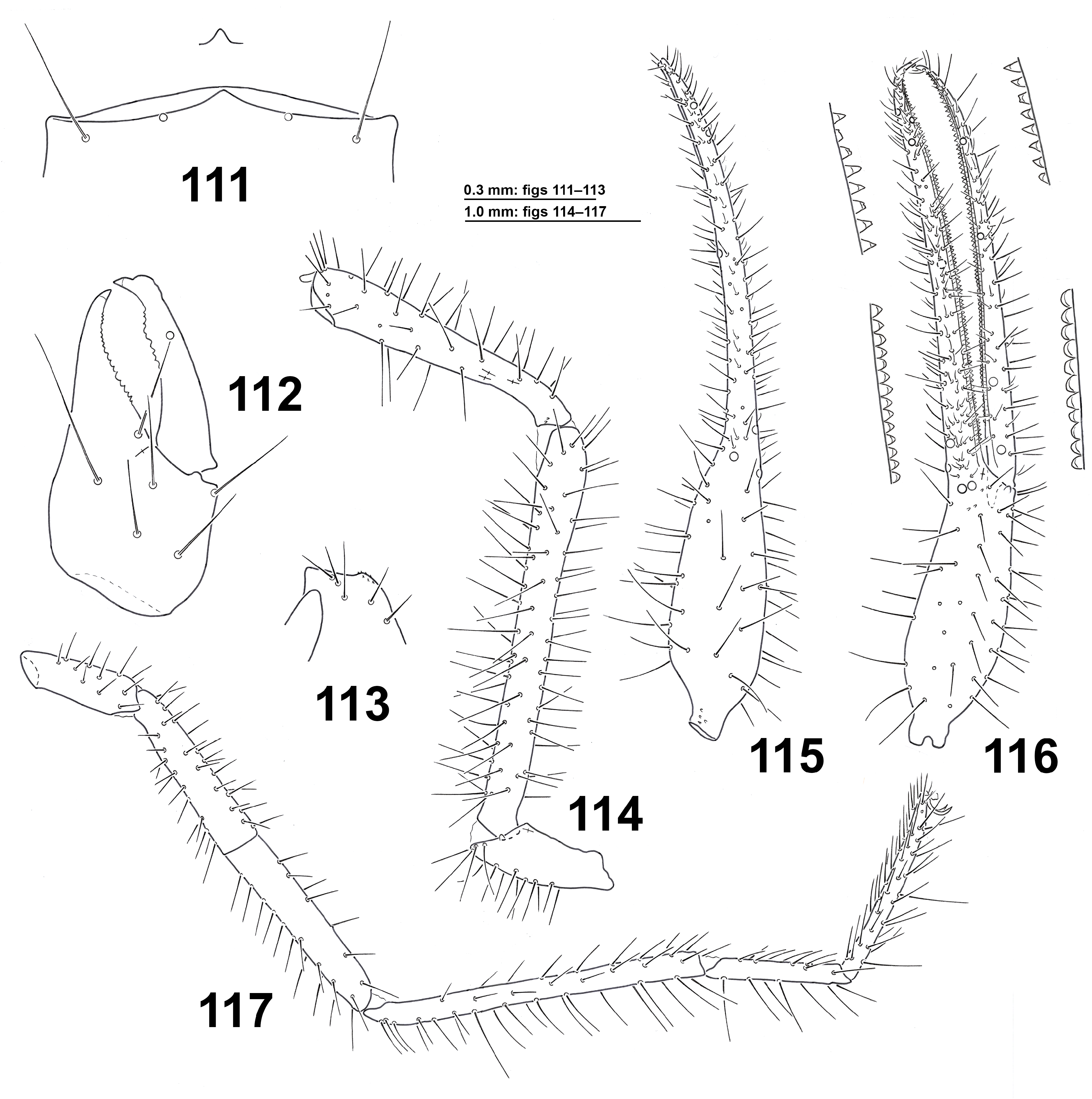
( Figs 111–118 View FIGURES 111–117 View FIGURE 118 )
Neobisium ( Ommatoblothrus) zoiai Gardini & Rizzerio, 1986: 6 View in CoL , figs 1–4.
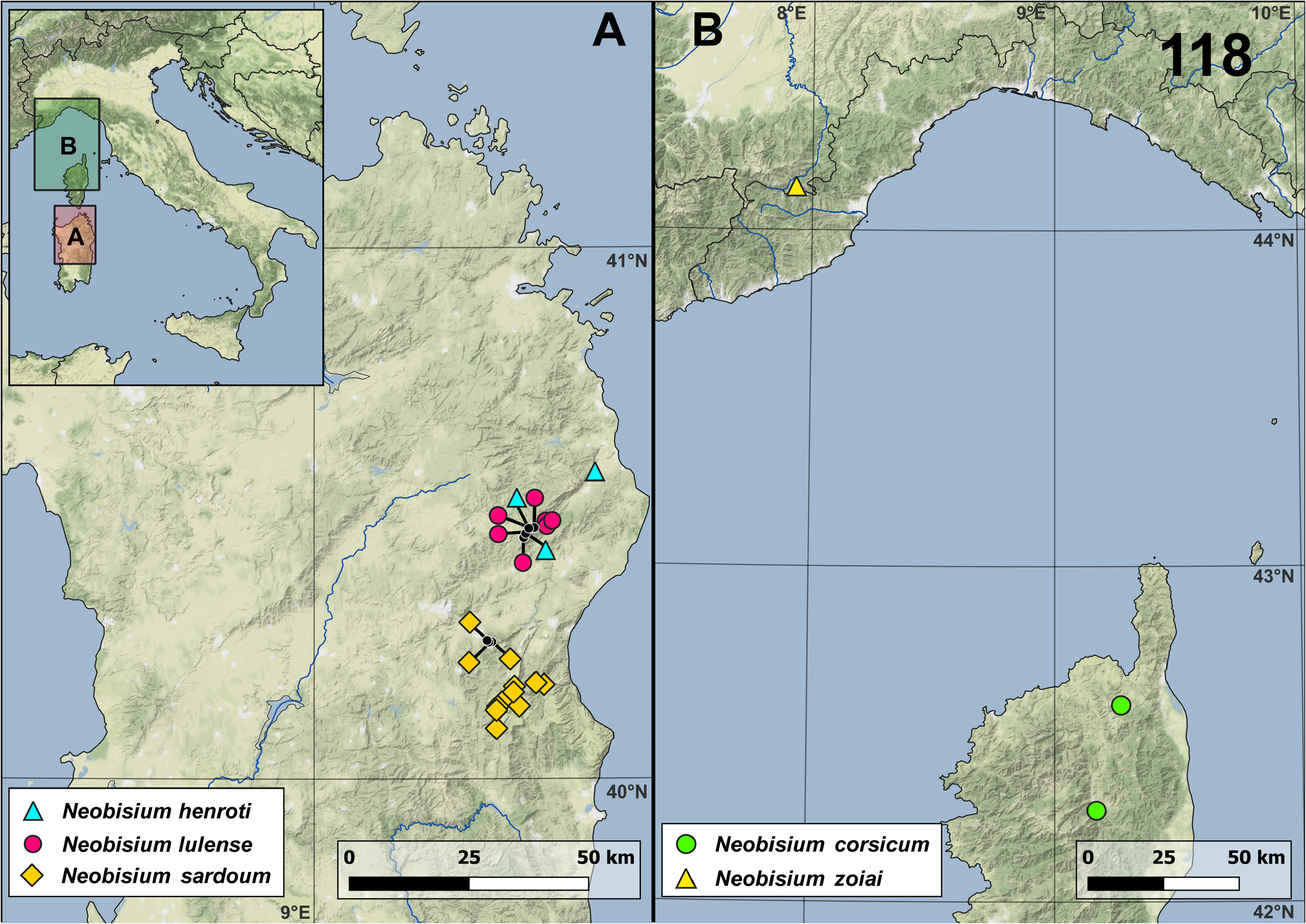
Type locality: Italy, Piedmont, Cuneo Province, Caprauna, Garb del Dighea 126 Pi / CN ( 40°07’45”N 7°56’00”E) GoogleMaps .
Distribution. Italy ( Piedmont).
Diagnosis ( ♂ ♀). A subterranean, highly troglomorphic Neobisium from Piedmont (Ligurian Alps) that differs from other Mediterranean species of the genus in the following combination of characters: posterior margin of carapace with 4 setae, preocular setae absent; epistome triangular or slightly prominent, anterior eyes weakly developed, with flat lens, dark integument between anterior and posterior eyes absent; tergites I–IV: 4–5:5–6:5– 6;6–7; cheliceral palm with 6 (rarely 7) setae, rallum with 9 (rarely 7) blades; fixed chelal finger with large, pointed, erected and spaced teeth in the distal three-quarters, more or less rounded in the proximal quarter of the finger; fixed and movable chelal fingers with 94–98 and 83–88 teeth, respectively; pedipalpal femur 2.32–2.46 mm length (8.43–8.94 ×), patella 1.70–1.88 mm (5.66–6.06 ×), movable finger 2.41–2.48 mm; pedipalpal hand with long oval profile, just widened towards the base and its greatest width in the proximal quarter; fixed finger curved in dorsal view, slightly curved, almost straight, in antiaxial view; ratio between movable chelal finger and hand of chela with pedicel 1.42–1.56; ratio between pedipalpal femur and movable chelal finger 0.96–1.00; ratio between pedipalpal femur and carapace 1.95–2.21; trichobothrium ist proximal st, at midway between the finger tip and isb.
Type material examined. ITALY — Piedmont: Cuneo Prov.: 1♀ ( holotype), Caprauna, Monte Armetta , Garb del Dighea 126 Pi / CN ( 40°07’45”N 7°56’00”E), 1615 m a.s.l., 17.VI.1985, G. Gardini, R. Rizzerio, S. Zoia leg. GoogleMaps
Non-type material examined. ITALY — Piedmont: Cuneo Prov.: 3♂, Caprauna, Monte Armetta , Garb del Dighea 126 Pi / CN ( 40°07’45”N 7°56’00”E), 1615 m a.s.l., 7.VIII.1994, G. Gardini, C. Giusto leg. GoogleMaps ; 1♀ 1 T, id., 2.II.2002, S. Zoia, F. Polese leg.
Description of adults ( ♂ ♀). Carapace, chelicerae, pedipalps and palpal coxae brown pale; pedipalps smooth, pleural membrane granular. Carapace 1.48 ( ♂) or 1.37–1.45 ( ♀) times as long as broad, anterior eyes weakly developed, with flat lens, the posterior ones not visible, dark integument between anterior and posterior eyes absent; anterior margin of carapace with a slightly prominent or triangular epistoma apically rounded ( Fig. 111 View FIGURES 111–117 ); 20 macrosetae, without preocular setae, chaetotaxy 4–6–6–4. Chaetotaxy of tergites I–X: 5:5–6:5–6:6–7:6– 7:8–11:10–11:8–11:10–11:10–11 ( ♂) or 4–5:5:6:6–7:7:7:8–9:9–10:11:10 ( ♀). Chaetotaxy of sternites II–X ( ♂): 22:(4)50(4):(3)16–17(3):15–17:15:15–16:15–16:15–16:13–14, median genital sac pyriform, reaching the base of sternite IV; chaetotaxy of sternites II–X ( ♀): 10:(4)23–24(4):(3)18(3):15–18:16–17:16:15–16:14–15:15. Chelicera ( Fig. 112 View FIGURES 111–117 ) 1.97–2.10 ( ♂ ♀) times as long as broad, palm with 6 setae (6 and 7 in one ♂); fixed finger with 17–23 subequal teeth proximally and distally decreasing in size; movable finger with 15–18, of which 1–3 more prominent at level of gs; gs ratio 0.64–0.68, spinneret largely prominent and rounded in male, more prominent in female ( Gardini & Rizzerio 1986, fig. 1); rallum with 9 blades, the distal one laterally pinnate and isolated on a prominence, the second one serrate, the successive ones smooth, the two proximal shorter ( one male with 7 and 9 blades in the right and left chelicera, respectively); serrula exterior with 35–38. Manducatory process with 5 setae. Coxal setae: pedipalp 6–8, I 5–8, II 6–10, III 6–8, IV 8–11; anterolateral process of coxa I broad and non-pointed, anteromedial process broadly rounded, with spicules ( Fig. 113 View FIGURES 111–117 ). Pedipalp ( Figs 114–116 View FIGURES 111–117 ): trochanter 3.05 ( ♂) or 2.83–2.92 ( ♀) times as long as broad; femur 8.43–8.94 ( ♂ ♀) times as long as broad, weakly enlarged distally; patella 5.66 ( ♂) or 5.77–6.06 ( ♀) times as long as broad, club gradually enlarged distally, ratio between club and pedicel 3.04 ( ♂) or 2.08–2.10 ( ♀), ratio X/Y = 0.25–0.27 ( ♂ ♀); chela with pedicel 7.60 ( ♂) or 7.36–7.46 ( ♀) times as long as broad; hand of chela with pedicel 3.02 ( ♂) or 3.05–3.11 ( ♀) times as long as broad, with long oval profile, just widened towards the base and its greatest width in the proximal quarter (dorsal view); fixed finger curved in dorsal view, slightly curved, almost straight, in antiaxial view ( Figs 115–116 View FIGURES 111–117 ; Gardini & Rizzerio 1986, figs 3–4); fixed chelal finger with 94–98 ( ♂ ♀) large, pointed, erected and spaced teeth in the distal three-quarters of the finger, more or less rounded in the proximal quarter of the finger ( Fig. 116 View FIGURES 111–117 ); nodus ramosus subterminal; movable chelal finger with 83–88 teeth, pointed, erected and spaced in the distal half, rounded and contiguous in the proximal half of the teeth ( Fig. 116 View FIGURES 111–117 ); all teeth with dental canals; sensillum closer to st than to sb; trichobothria as in figs 115–116, relative position of trichobothria along chelal axis ( ♂ ♀): it 0.11/ et 0.13–0.15/ est 0.145 –0.18/ ist 0.44–0.47/ isb 0.85–0.88/ ib 0.93–0.94/ esb 0.98/ eb 0.99/ t 0.19–0.20/ st 0.34–0.42/ sb 0.75–0.81/ b 0.90–0.91; trichobothrium ist proximal st, at midway between the finger tip and isb; trichobothrium isb proximal sb; ratio between movable finger and hand of chela with pedicel 1.56 ( ♂) or 1.42–1.46 ( ♀); ratio between pedipalpal femur and movable finger 0.96–1.0 ( ♂ ♀); ratio between pedipalpal femur and carapace 1.95 ( ♂) or 2.06–2.21 ( ♀). Leg IV ( Fig. 117 View FIGURES 111–117 ): trochanter 3.09 ( ♂) or 2.69–3.26 ( ♀) times as long as deep, femur + patella 9.32 ( ♂) or 8.61–10.08 ( ♀) times as long as deep, tibia 13.44 ( ♂) or 14.41–15.57 ( ♀) times [TS = 0.28–0.31 ( ♂ ♀)], basitarsus 6.71 ( ♂) or 7.42–8.0 ( ♀) times [TS = 0.11–0.14 ( ♂ ♀)], telotarsus 11.5 ( ♂) or 9.58–10.45 ( ♀) times as long as deep [TS = 0.40–0.50 ( ♂ ♀)], ratio between basitarsus and telotarsus ( ♂ ♀) 0.70–0.80, subterminal seta furcate, claws without dorsal tooth.
Measurements. Body length 4.8 ( ♂) or 5.0–5.4 ( ♀). Carapace 1.19 × 0.80 anteriorly ( ♂) or 1.11–1.19 × 0.81– 0.82 anteriorly ( ♀). Chelicera 0.73 × 0.37 ( ♂) or 0.80–0.815 × 0.38–0.39 ( ♀); movable finger length 0.46 ( ♂) or 0.505–0.51 ( ♀). Pedipalp: trochanter 0.84 × 0.275 ( ♂) or 0.805–0.82 × 0.275–0.29 ( ♀); femur 2.32 × 0.275 ( ♂) or 2.46 × 0.275–0.29 ( ♀); patella 1.70 × 0.30 ( ♂) or 1.79–1.88 × 0.31 ( ♀); chela with pedicel 3.88 × 0.51 (depth 0.515) ( ♂) or 4.05–4.18 × 0.55–0.56 (depth 0.55–0.56) ( ♀); hand with pedicel length 1.54 ( ♂) or 1.68–1.74 ( ♀); movable finger length 2.41 ( ♂) or 2.46–2.48 ( ♀). Leg IV: trochanter 0.725 × 0.235 ( ♂) or 0.70–0.75 × 0.23–0.26 ( ♀); femur + patella 2.19 × 0.235 ( ♂) or 2.32–2.41 × 0.23–0.28 ( ♀); tibia 1.95 × 0.145 ( ♂) or 2.09–2.18 × 0.14–0.145 ( ♀); basitarsus 0.805 × 0.12 ( ♂) or 0.89–0.92 × 0.115–0.12 ( ♀); telotarsus 1.15 × 0.10 ( ♂) or 1.15 × 0.11–0.12 ( ♀).
Description of tritonymph. Carapace, chelicerae, pedipalps and palpal coxae brown pale. Pedipalps smooth, pleural membrane granular. Carapace 1.39 times as long as broad, eyes or eye spots not visible; anterior margin of carapace with slightly prominent, broadly rounded epistome; carapace with 20 macrosetae, without preocular setae, chaetotaxy 4–5–7–4. Chaetotaxy of tergites I–X 4:5:5:8:9:8:8:11:11:9. Chaetotaxy of sternites II–X: 4:(2)11(2):(3)11(3):15:14:15:13:15:12. Chelicera 2.13 times as long as broad, palm with 6 setae, fixed finger with 21 teeth, movable finger with 16 teeth, of which two prominent at level of gs; gs ratio 0.66, spinneret largely prominent, hemispheric; rallum with 7 blades, the distal one laterally pinnate and isolated on a prominence, the second and the third ones distally serrate, the successive ones smooth, the proximal shorter; serrula exterior with about 30 blades. Manducatory process with 4 setae. Coxal setae: pedipalp 6, I 5, II 6, III 5, IV 8; anterolateral process of coxa I short and stocky. Pedipalp: trochanter 2.78 times as long as broad; femur 6.62 times as long as broad, weakly enlarged distally; patella 4.61 times as long as broad, weakly and gradually enlarged distally, club 2.0 times longer than pedicel, ratio X/Y = 0.35; chela with pedicel 6.34 times as long as broad; hand of chela with pedicel 2.70 times as long as broad; fixed chelal finger with 70 teeth: 3 small, rounded distal teeth, followed by 47 large, pointed, erected and spaced teeth and 20 rounded teeth increasingly reduced towards finger base; nodus ramosus subterminal; movable chelal finger with 62 teeth, pointed and erected in the distal half of the teeth, rounded in the proximal one; all teeth with dental canals; sensillum not seen; relative position of trichobothria along chelal axis: it 0.13/ et 0.16/ est 0.21 / ist 0.50/ ib 0.93/ esb 0.98/ eb 0.985/ t 0.22/ st 0.50/ b 0.88; ratio between movable finger and hand of chela with pedicel 1.43; ratio between pedipalpal femur and movable finger 0.93; ratio between pedipalpal femur and carapace 1.72. Leg IV: trochanter 2.63 times as long as deep, femur + patella 7.78 times as long as deep, tibia 10.58 times (TS = 0.34), basitarsus 5.6 times (TS = 0.13), telotarsus 7.27 times as long as deep, ratio between basitarsus and telotarsus 0.70, subterminal seta furcate, with short dorsal branch; claws without dorsal tooth.
Measurements. Body length not detected for opisthosoma broken. Carapace 0.92 × 0.66 anteriorly. Chelicera 0.64 × 0.30, movable finger length 0.38. Pedipalp: trochanter 0.64 × 0.23; femur 1.59 × 0.24; patella 1.20 × 0.26; chela with pedicel 2.79 × 0.44 (depth 0.44); hand with pedicel length 1.19; movable finger length 1.71. Leg IV: trochanter 0.50 × 0.19; femur + patella 1.48 × 0.19; tibia 1.27 × 0.12; basitarsus 0.56 × 0.10; telotarsus 0.80 × 0.11.
Remarks. Neobisium zoiai was described by Gardini & Rizzerio (1986) based on one female from the cave Garb del Dighea 126 Pi/CN near Caprauna (Ligurian Alps), so far the only locality known for the species. It is a relictual species of great zoogeographic interest, which is geographically isolated and with a pattern of chelal fingers dentition that differentiates it from all the other subterranean species of Neobisium from the Corso-Sardinian complex and from central and southern Italy. The proximal position of the trichobothrium ist, almost in the middle of the fixed chelal finger, is also unique among the subterranean Neobisium species studied here.
Neobisium zoiai was compared by Gardini & Rizzerio (1986) with Neobisium gomezi Hertault, 1979 , described on a male from a cave near Sainte-Engrâce, Pyrénées Atlantiques ( France), which shares with N. zoiai a similar dentition, the absence of preocular setae, four setae on posterior margin of carapace and the same position of trichobothrium ist.
Evaluation of the variability of some characters currently used in the taxonomy of the genus Neobisium
The examination of specimens of Neobisium previously attributed to the subgenus Ommatoblothrus ―the subject of this revision―has made it possible to evaluate the variability of some characters traditionally used in taxonomy. Only the variability present in antimere appendages of single specimens or in specimens of the same population is taken into consideration, and morphometric characters are excluded. The species that have allowed a greater evaluation of the chaetotaxic characters are N. leolatellai n. sp. and N. petruzzielloi n. sp., of which numerous specimens have been available for each population.
Eyes. It is not easy to objectively express the level of reduction of the eyes, expressed for the anterior ones― rarely reduced to eyespots―by the greater or lesser convexity of the lens, and for the posteriors by the presence of a flat lens down to a small eyespot. The population of N. leolatellai n. sp. from the Grotta della Cava is the only one to show a certain variability in this regard, with the lens of the anterior eyes ranging from convex to almost flat. The presence of tapetum remains in single males of N. leolatellai n. sp. from the Grotta della Cava and of N. petruzzielloi n. sp. from the Grotta del Caprone needs to be confirmed; however, the presence of a weak tapetum was mentioned by Beier (1956b) for N. sardoum .
Setae on posterior margin of carapace. It varies from 4 to 8 but, depending on the species considered, the standard conditions are either 4 or 6, perhaps 8 only in N. lulense . Four (rarely 5) posterior setae are present in adults of N. leolatellai n. sp. from Grotta della Cava and Grotta di Fiume Coperto, N. magrinii n. sp. from Grotta dell’Arnale, N. vignai n. sp. from Grotta degli Ulivi and N. petruzzielloi n. sp. from Grotta degli Sportiglioni, but N. petruzzielloi n. sp. from Grotta del Caprone and Grotta dei Diavoli have 4 or 5 or 6 setae equally distributed among specimens, so it is not possible to hypothesize which number prevails in these populations.
Chaetotaxy of tergites I–IV. Tergal chaetotaxy usually shows a high intrapopulational variability in Neobisium species, and that of the first tergites often repeats the same number of setae as the posterior margin of carapace. Neobisium leolatellai n. sp. from Grotta della Cava and Grotta di Fiume Coperto has tergite I mostly with 4 setae (in 12 adults, 2 tritonymphs and 1 protonymph), and tergites I–IV with chaetotaxy 4–4–4– 4 in 6 adults and 2 tritonymphs; also, 2 adults have 2 setae on tergite I, 3 adults, 2 tritonymphs and 1 deutonymph have 3 setae on tergite I. Tergal chaetotaxy in adults of N. petruzzielloi n. sp. is more variable, but it is presumable that the standard one of tergites I–IV of two of the known populations (Grotta degli Sportiglioni and Grotta del Caprone) is 6–6–6–6. In the 28 adult specimens examined, the number of setae in tergites I-IV is as follows: tergite I: 6 specimens with 4 setae, 9 with 5, 11 with 6, 1 with 7 and 1 with 8; tergite II: 2 specimens with 5 setae, 22 with 6, 1 with 7 and 3 with 8; tergite III: 21 specimens with 6 setae, 7 with 7; tergite IV: 8 specimens with 6 setae, 11 with 7, 7 with 8 and 2 with 9. In the case of N. petruzzielloi n. sp. the supposed standard number of 6 setae on tergite I does not correspond to that of the posterior margin of carapace (4 setae).
Chaetotaxy of sternites II–III. The number of setae present on sternites II and III appears to be highly variable, especially in males, within the same population. Neobisium leolatellai n. sp. has sternite II with 17–23 setae and sternite III with 38-50 setae in males from Grotta della Cava. Females of the same species have lower variability: sternite II with 9–13 setae and sternite III with 23–30 setae (Grotta della Cava, Grotta di Fiume Coperto and Grotta del Laghetto Sulfureo). The validity of these characters as interspecific discriminants needs to be further evaluated.
Setae on manducatory processes. Almost all specimens of Neobisium examined have the same number of setae on both the manducatory processes: 5 in adults, 4 in tritonymphs and 3 in deutonymphs. Few specimens of 4 species have 4 and 5 setae, a female of N. leolatellai n. sp. from Grotta di Fiume Coperto has 4 and 4 setae and the two known females of N. corsicum n. sp. have 3 and 4 and 4 and 4 setae, respectively.
Coxal setae. The variability of coxal chaetotaxy is so high―both within the single specimens and within the same population―that it seems impossible to find a standard formula for each species. Undoubtedly, it would be necessary to examine a greater number of specimens for each population.
Cheliceral teeth. See Remarks under N. sardoum .
Setae on cheliceral palm. The number of setae present on the cheliceral palm was used in the past as a differential character to discriminate even groups of species in the genus Neobisium (see Beier 1963: 117), but its use as a specific discriminant was strongly discouraged by Vachon (1966). Six setae are normally present on both cheliceral palms of species studied here, with a few exceptions: 5 and 6 in one female of N. leolatellai n. sp. (Cave towards the Madonna della Guardia), 6 and 7 in one male of N. patrizii (Grotta Pozzo l’Arcaro) and in one male of N. zoiai (Garb del Dighea). Specimens of N. vignai n. sp. from Grotta degli Ulivi have 6 and 6 setae ( 2♀), 6 and 7 setae ( 2♀) and 7 and 7 setae ( 1♀), and so N. petruzzielloi n. sp. from Grotta del Caprone has 6 and 6 setae ( 1♂ 1♀), 6 and 7 setae ( 1♂ 1♀) and 7 and 7 setae ( 3♂ 5♀). Specimens of N. oenotricum have 7 and 7 setae, with the exception of the male holotype, with 7 and 8 setae ( Callaini 1987).
Cheliceral rallum. The importance of the structure of the rallum in the taxonomy at the genus level within the Neobisiidae has been known for some time (Vachon 1964; Vachon & Gabbutt 1964; Mahnert 1974). The number of rallar blades, together with their mutual position and structure (pinnate or denticulate/serrate or smooth), are characters currently used in the discrimination of the genera of the subfamily Neobisiinae ( Nassirkhani et al. 2019) . The standard model of rallum of the Neobisium species coincides with that found by Mahnert (1988) for adults of N. carcinoides : 8 blades, of which the distal one is pinnate/serrate and isolated on a prominence, the second one serrate, the next five smooth and gradually shorter, the proximal one very short and often isolated from the others. This condition is present in 47% of the adults examined. The tritonymphs and deutonymphs have the same structure as the adults, but with a smaller number of blades: 7 and 6, respectively. Variations on the standard model of the rallum have already been highlighted above (see Introduction, with assessments of the taxonomic status of the subgenera of Neobisium Chamberlin, 1930 ). The populations of N. leolatellai n. sp. from Lepini Mountains are those with the highest number of blades and the greatest variability: 10–13 blades, of which 1 distal pinnate, 1 serrate, 5–7 smooth and 2–3 smooth very short, in adults; 7–9 blades, of which 1 distal pinnate/serrate, 1–2 serrate, 5–7 smooth (of which 1 or 2 proximal very short), in tritonymphs; 7–8 blades, of which 1 distal pinnate/serrate, 1 serrate, 5–6 smooth (of which 1 or 2 proximal very short), in deutonymphs.
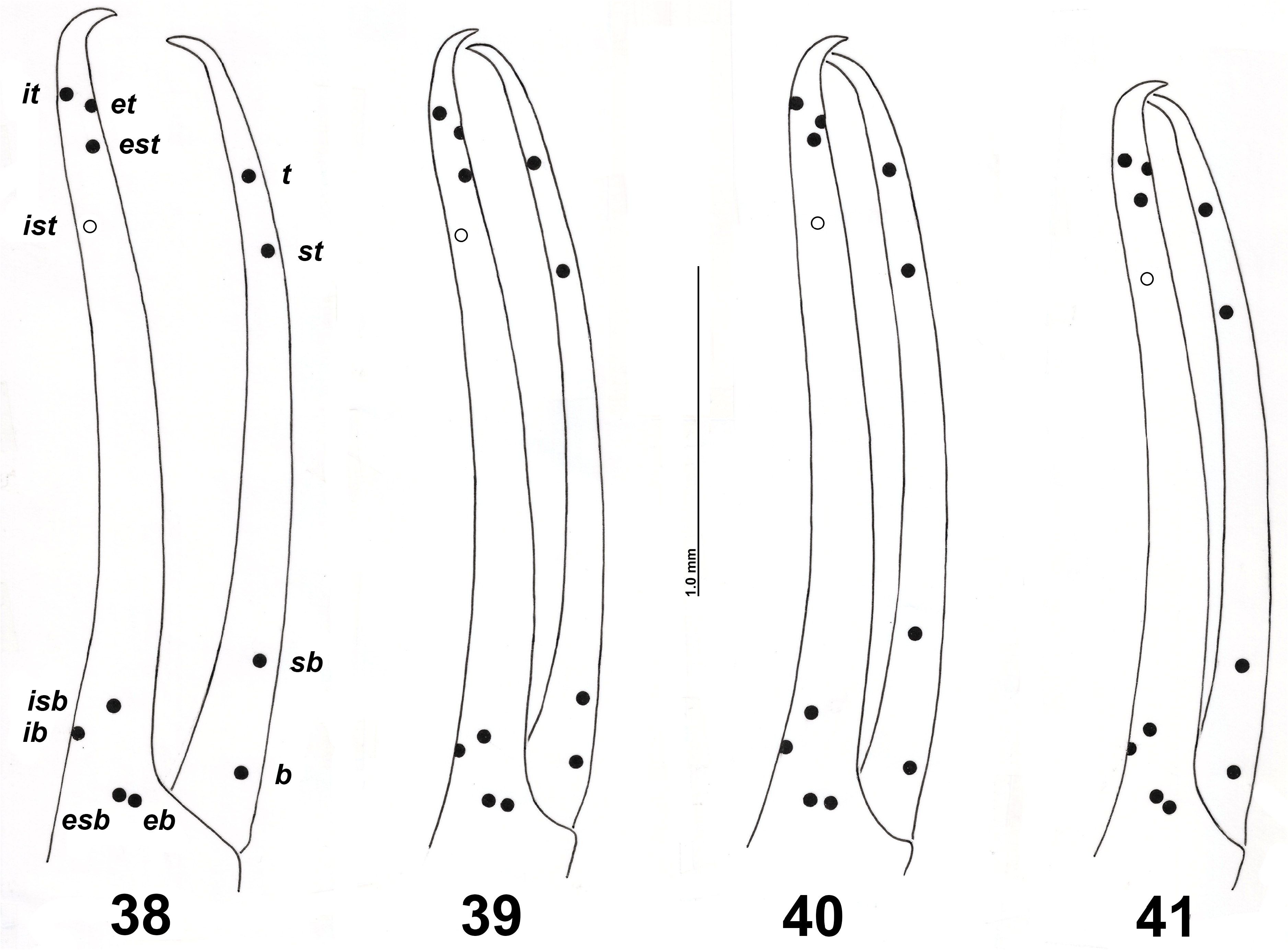
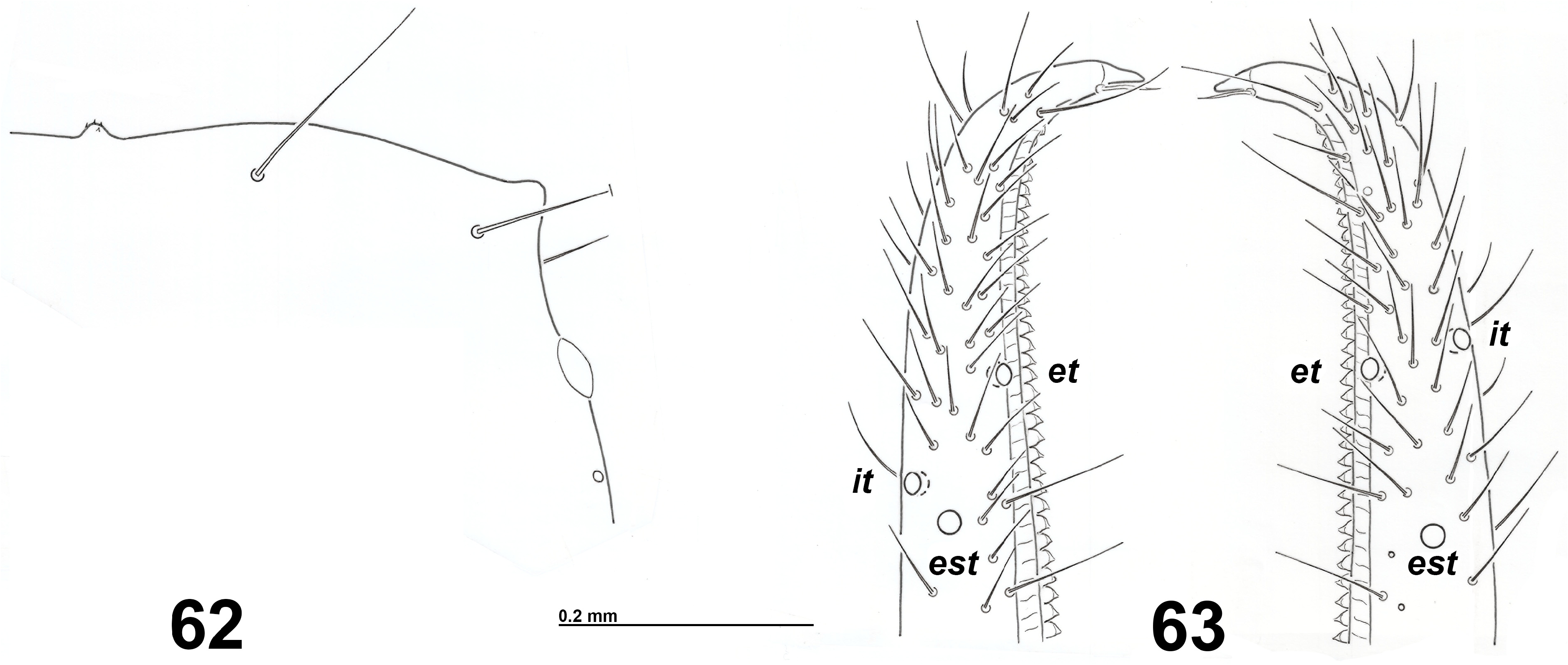
Trichobothria. The positions of the trichobothria on both pedipalpal fingers provide important characters in the systematics at the supraspecific and generic level of neobisiids. But their importance as specific discriminants within the genus Neobisium has often been overestimated, as the intraspecific variability was not considered, often due to the scarcity of specimens available for study. In the descriptions of Neobisium species, and in the relative keys, the mutual positions of the following trichobothria in adults are mainly considered ( Beier 1963; Vachon 1976; Heurtault 1986; Mahnert 1988; Novák 2014; Nassirkhani et al. 2020): i) the position of the group it-et-est (or only of est) with respect to t; ii) the position of ist with respect to st; iii) the position of ist with respect to it, or to the apex of the fixed finger, and to ib (i.e., the position of ist on the fixed finger). The mutual positions of it-et-est varies even within the same population ( Figs 39–40 View FIGURES 38–41 and Mahnert 1977, figs 34a–b–c), rarely in the same specimen ( Fig. 63 View FIGURES 62–63 ). With reference to the species covered in this review, the position of est can vary from proximal to the same level with respect to t in N. sardoum (Grotta Sa Nurra de Sas Palumbas and Grotta Su Conti) , N. magrinii n. sp. (Grotta l’Arnale), N. petruzzielloi n. sp. (Grotta del Caprone) and N. vignai n. sp. (Grotta degli Ulivi), or the position of est can vary more widely from proximal to distal with respect to t in N. leolatellai n. sp. (Grotta della Cava), N. oenotricum (Grotta Sant’Angelo) and N. sp. aff. patrizii (Grotta l’Arnara) . As for the position of ist relative to st, the position of ist can vary from proximal to the same level with respect to st in N. sardoum (Grotta Sa Nurra de Sas Palumbas) and N. petruzzielloi n. sp. (Grotta degli Sportiglioni), or the position of ist can vary more widely from proximal to distal with respect to st in N. leolatellai n. sp. (Grotta della Cava, Grotta di Fiume Coperto and Grotta del Laghetto Sulfureo), N. magrinii n. sp. (Grotta l’Arnale) and N. petruzzielloi n. sp. (Grotta del Caprone). The relative position of trichobothrium ist along the chelal axis varies from 0.23 to 0.42 in the Neobisium species reviewed here, apart from N. zoiai , for which it is 0.44–0.47. Intrapopulational variability is mostly low: 0.28–0.32 in N. leolatellai n. sp. (Grotta della Cava) and 0.315–0.35 in N. petruzzielloi n. sp. (Grotta degli Sportiglioni and Grotta del Caprone).
Subterminal setae of telotarsi of legs I–IV. Chamberlin (1931:163 and fig. 40 O–T) states that the subterminal setae of the telotarsi have a general systematic interest and that “forked subterminal setae are almost confined to the Neobisiidae ”. From Chamberlin (1931) to the present, the shape of the subterminal setae of the telotarsi of Neobisiidae , mostly confined to leg IV, has been the subject of some taxonomic interest at species level. The subterminal setae of telotarsi on both sides of legs I-IV of a randomly selected male of N. sp. aff. patrizii from the Grotta dell’Arnara are represented in fig. 8. The high variability of the shape of the subterminal setae of telotarsi, even in a single specimen, clearly shows its scarce utility as a specific discriminant.
| CN |
Wellcome Collection of Bacteria, Burroughs Wellcome Research Laboratories |
| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Neobisium |
Neobisium ( Neobisium ) zoiai Gardini & Rizzerio, 1986
| Gardini, Giulio 2023 |
Neobisium ( Ommatoblothrus ) zoiai
| Gardini, G. & Rizzerio, R. 1986: 6 |