
Myriapora bugei d’Hondt, 1975
|
publication ID |
https://doi.org/ 10.5852/ejt.2013.44 |
|
publication LSID |
lsid:zoobank.org:pub:F7FD3319-AD9D-4DBB-9755-C541759C0D66 |
|
DOI |
https://doi.org/10.5281/zenodo.3815230 |
|
persistent identifier |
https://treatment.plazi.org/id/324587E8-FF86-D83A-8A43-FA58FE874E39 |
|
treatment provided by |
Carolina |
|
scientific name |
Myriapora bugei d’Hondt, 1975 |
| status |
|
Myriapora bugei d’Hondt, 1975 View in CoL
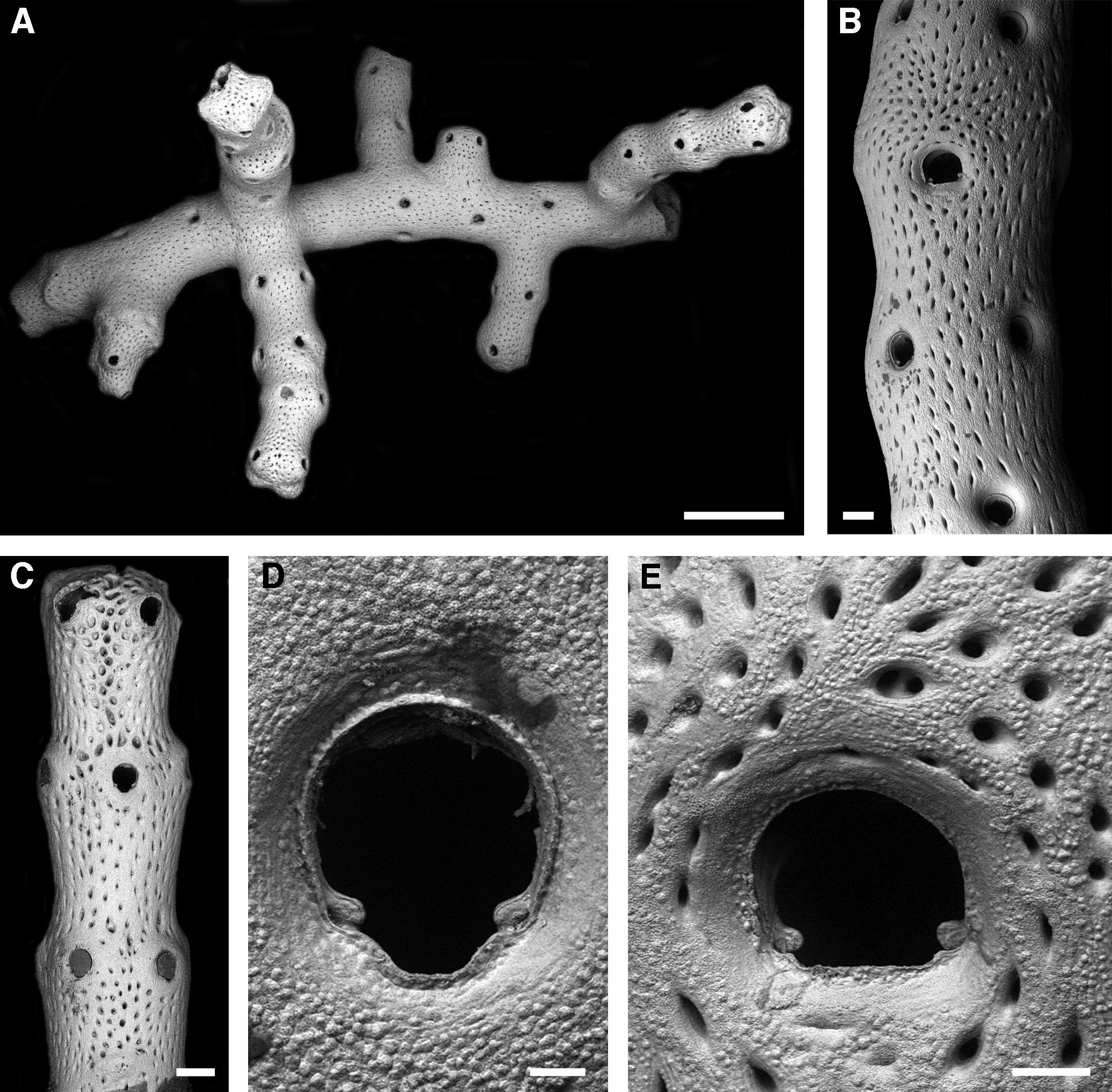
Fig. 4 View Fig , Table 4
Myriapora bugei d’Hondt, 1975: 585 View in CoL , figs 23, 25-28.
non Myriapora bugei View in CoL – El Hajjaji 1992: 250, pl. 15, figs 14-15.
Material examined
Lectotype (here designated)
MNHN IB-2013-3 , one dried colony fragment (former part of MNHN 7481 About MNHN ), Jean Charcot GoogleMaps , Biaçores Stn 109, 20 Oct. 1971, NW of Flores (Azores), 39°33’ N – 31°17’ W, 190–230 m.
Paralectotypes (here designated)
MNHN 7481, Jean Charcot, Biaçores Stn 109, 20 Oct. 1971, NW of Flores (Azores), 39°33’ N – 31°17’ W, 190–230 m; MNHN IB- 2013-2, one dried colony fragment (former part of MNHN 7492), Jean Charcot, Biaçores Stn 110, 20 Oct. 1971, NW of Flores (Azores), 39°33’ N – 31°17.5’ W, 300–350 m; MNHN 7482, 13 colony fragments in ethanol, Jean Charcot, Biaçores Stn 109, 20 Oct. 1971, NW of Flores (Azores), 39°33’ N – 31°17’ W, 190–230 m; MNHN 7488, one colony fragment in ethanol, Jean Charcot, Biaçores Stn 110, 20 Oct. 1971, NW of Flores (Azores), 39°33’ N – 31°17.5’ W, 300–350 m; MNHN 7492, four colony fragments in ethanol, Jean Charcot, Biaçores Stn 110, 20 Oct. 1971, NW of Flores (Azores), 39°33’ N – 31°17.5’ W, 300– 350 m.
The original material available to d’Hondt (1975: 560) was from Biaçores stations 109 and 110. However, he did not indicate type material when introducing Myriapora bugei , and the figured specimen (figs 23, 25-28) was subsequently destroyed (J.-L. d’Hondt pers. comm. 2012). For the same reasons as stated in the Material section for Cellaria harmelini (see above), the type specimens of M. bugei were invalidly considered as “ holotype ” and “ paratypes ” by Tricart & d’Hondt (2009), and are formally designated here as lectotype and paralectotypes, respectively.
Description
Colony erect, rigid, delicate branching, producing lateral offsets at an angle of about 90° at irregular intervals ( Fig. 4A View Fig ). Branches long (3-4 cm), slender and cylindrical, with the zooids opening all around. Zooids large, radially arranged in distinct whorls of 4 (rarely 5), with all orifices at same level and the distal whorl ideally alternating ( Fig. 4 View Fig B-C); zooecia elongate rectangular but boundaries invisible on colony surface. Frontal shield of mature zooecia gently raising towards orifice, giving distal branches an undulating morphology that is slightly reduced due to frontal calcification during ontogeny ( Fig. 4 View Fig A- C); frontal shield regularly perforated by numerous elongate pores in spindle-shaped depressions that are longitudinally aligned ( Fig. 4B View Fig ), surface finely granular ( Fig. 4 View Fig D-E). Primary orifice dimorphic; in autozooids slightly longer than wide ( Fig. 4D View Fig ), anter horseshoe-shaped, widest slightly distal to middistance, proximolateral corners rounded with the short shoulders slightly sloping towards a deep and broadly U-shaped sinus occupying about two-thirds of total proximal width, condyles conspicuous, broad and with rounded edges, not extending beyond shoulders of the proximolateral margin; orifice in ovicellate zooids D-shaped ( Fig. 4E View Fig ), of similar length as in autozooids but especially the proximal margin distinctly broader, sinus a very shallow straight edge comprising four-fifths of total proximal width, proximolateral shoulders thus relatively short and fairly straight, condyles similar to those in autozooids; orifice becoming immersed by frontal calcification and occasionally closed during ontogeny in proximal branch regions, in which case a small short peristome with a central pore remains. No oral spines.
Ovicells relatively rare, present in all zooids of certain whorls, indicated by a slightly thicker swelling of this branch region and by frontal pseudopores that are radially arranged from the proximal ooecium centre ( Fig. 4B, E View Fig ), ooecium mostly incorporated into frontal shields of the distal zooids, surface as frontal shield of zooecia, boundaries invisible at colony surface, ovicell aperture at an acute angle to frontal plane, closure of the cleithral or subcleithral type.
No avicularia.
An ancestrula was not present in the available material.
Remarks
Although the genus Myriapora comprises only a few species globally, owing to their mostly erect and conspicuous colonies, and because of the ubiquitous Mediterranean species M. truncata (Pallas, 1766) , this bryozoan genus has received a fair amount of attention (e.g., Viskova 1986; Berning 2007; Ferretti et al. 2007; Rodolfo-Metalpa et al. 2010). Besides M. truncata and the polar M. subgracilis (d’Orbigny, 1852) , M. bugei is the only other known species from the northern Atlantic realm. The most obvious difference between M. bugei and M. truncata is that in the former the branches are composed of distinct whorls whereas zooecia in the latter are ideally alternating, i.e., apparently arranged in spirals around the branch axis. The ooecia and dimorphic orifices of M. bugei are here described and figured for the first time.
Originally described from the Azores, the species has also been recorded from the Great Meteor Bank ( Piepenburg & Müller 2004: 61), which is located some 800 km south of the archipelago. However, although the characters are variable and overlapping to some extent, the populations from the Great Meteor Bank, as well as from the nearby Hyères and Irving seamounts, slightly differ from the type of M. bugei in having branches that predominantly consist of five zooids per whorl (occasionally four or six), smaller autozooids, and a narrower autozooecial sinus (pers. observation). Thus, especially considering the distance between the Azores and the seamounts, it is likely that these populations represent distinct species, although genetic studies may be needed to clarify this issue.
In the Azores, M. bugei was reported from depths between 190 and 1235 m ( d’Hondt 1975). It occurs on rocky, gravelly, sandy and muddy substrata growing on rocks, dead corals and bivalve shells.
The specimens El Hajjaji (1992) described and figured as M. bugei from the Late Miocene of NE Morocco (Mediterranean Sea) are similar to the type but certainly belong to a different species due to a distinctly narrower sinus and larger condyles. The status of other coeval specimens from NW Morocco (Atlantic) cannot be assessed, as a detailed description or figures were not provided by Sefian et al. (1999: 242).
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Myriapora bugei d’Hondt, 1975
| Berning, Björn 2013 |
Myriapora bugei
| El Hajjaji K. 1992: 250 |
Myriapora bugei d’Hondt, 1975: 585
| d'Hondt J. - L. 1975: 585 |