
Metaprotella africana Mayer, 1903
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4098.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:A041BA37-A806-4726-BD1C-92FEF2280F62 |
|
DOI |
https://doi.org/10.5281/zenodo.5195996 |
|
persistent identifier |
https://treatment.plazi.org/id/177687D7-FFE6-FFA5-E0CB-FF0CD340FC3D |
|
treatment provided by |
Plazi |
|
scientific name |
Metaprotella africana Mayer, 1903 |
| status |
|
Metaprotella africana Mayer, 1903
( Figs. 4–7 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Metaprotella africana Mayer, 1903: 42 ,43, pl. 1, figs. 32, 33; Ruffo, 1959: 23.
Material examined. Types, 1 male MNHN Am -5252 and 1 female MNHN Am -5253, collected from Djibouti, Somaliland (see Mayer, 1903)
Description. Male (MNHN Am -5252)
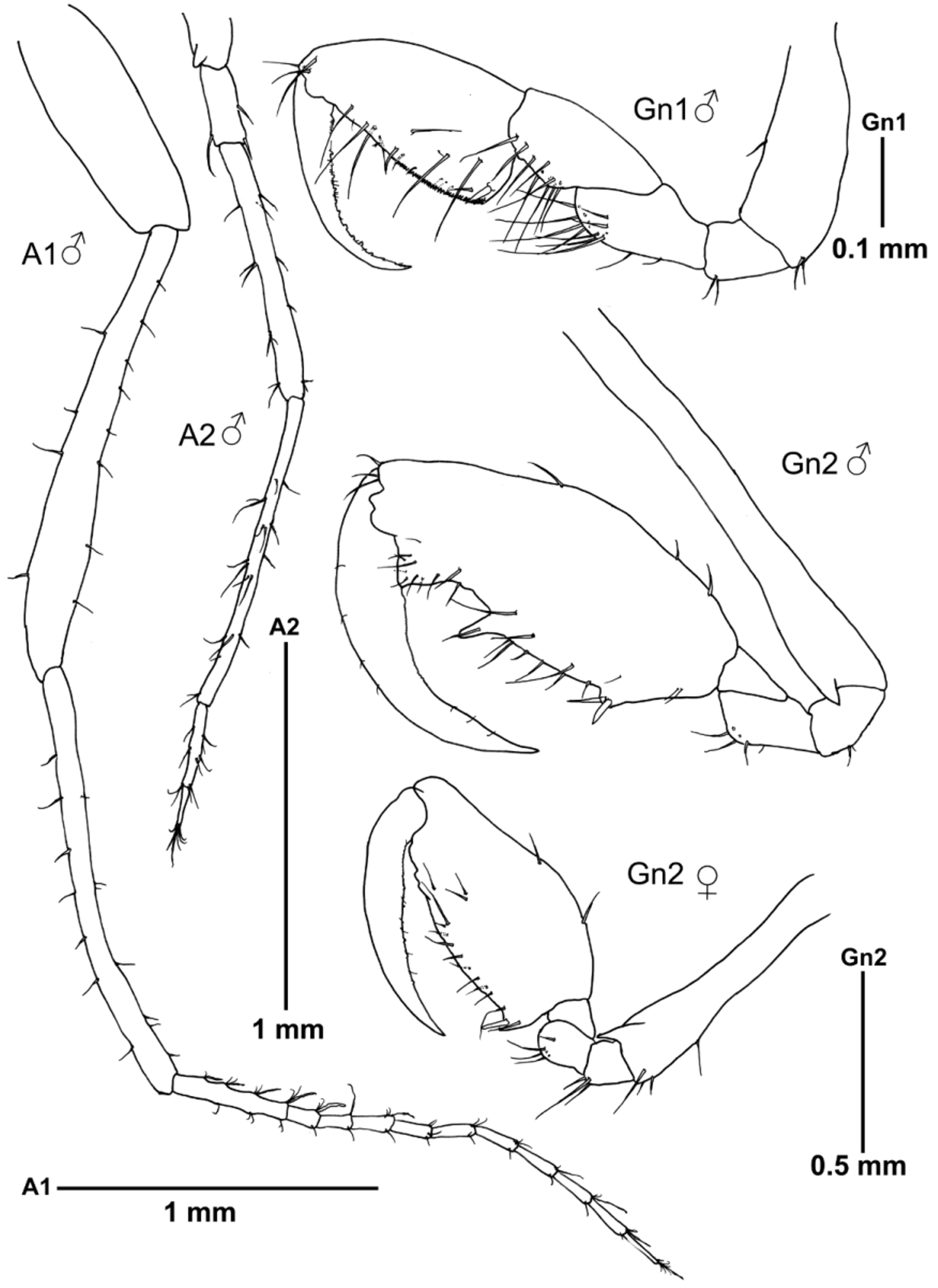
Body length 7.4 mm. Suture between head and pereonite 1 present; pereonite 3 longest. Pereonites 6 and 7 fused. Head and pereonites smooth, except for pereonite 2 with paired of anteriorly curved dorsal projections medially, and two small acute projections laterally near the coxa of gnathopod 2 ( Fig. 4 View FIGURE 4 A). Eye distinctive. Gills on pereonites 3 and 4, elongate, length about 3 times the width; first pair 1.2 times as long as the second pair.
Mouthparts. Upper lip symmetrically bilobed; each lobe carrying a distal row of minute setulae. Mandibles with palp (broken in left mandible); mandibular molar process strong; left mandible with incisor and lacinia mobilis divided into 5 teeth, followed by 3 submarginal pectinate setae; right mandible incisor divided into 5–6 teeth followed by lacinia mobilis divided into 3 teeth, one wider and minutely serrate, followed by 2 pectinate setae; palp with 3 articles; article 1 lacking setae; article 2 with 7 setae; article 3 with setal formula 1-4-1 and distal row of minute setulae. Inner lobes of the lower lip well demarcated, oval; outer and inner lobes provided with setulae. Maxilla 1 outer lobe carrying 7 bifurcate spines; distal article of palp with 3 teeth and 3 spines on apical end, and a group of setae medially. Maxilla 2 outer lobe rectangular, carrying 15 setae; inner lobe oval, with group of setae distally and setulae laterally. Maxilliped inner plate with 3 plumose setae and 2 non plumose setae on distal margin; outer plate larger than inner, 2 x length of inner plate, margin setulose, bearing 1 seta apically; palp setose, dactylus falcate, with two rows of setulae and a proximal seta on outer margin.
Antenna 1 about 0.7 x body length; article 2 of the peduncle the longest; flagellum 11-articulate, with the proximal article composed of 4 articles fused. Antenna 2 slender, about 0.4 x antenna 1 length, without swimming setae; peduncular article 1 with a short acute distal projection; flagellum 2-articulate.
Gnathopod 1 basis shorter than ischium to carpus combined; propodus oval with proximal grasping spine, palm margin finely serrate; dactylus minutely denticulate and setose on inner margin. Gnathopod 2 inserted on the middle of pereonite 2; coxa vestigial, small acute projection near the coxa, basis slender, 1.2 x pereonite 2 length, with a small acute projection distally; propodus elongate, length about 2.3 times of width, with a grasping spine on a small proximal projection, medial projection, followed by U -notch and triangular projection distally; dactylus with sparse setulae and minutely denticulate in the proximal and medium margin.
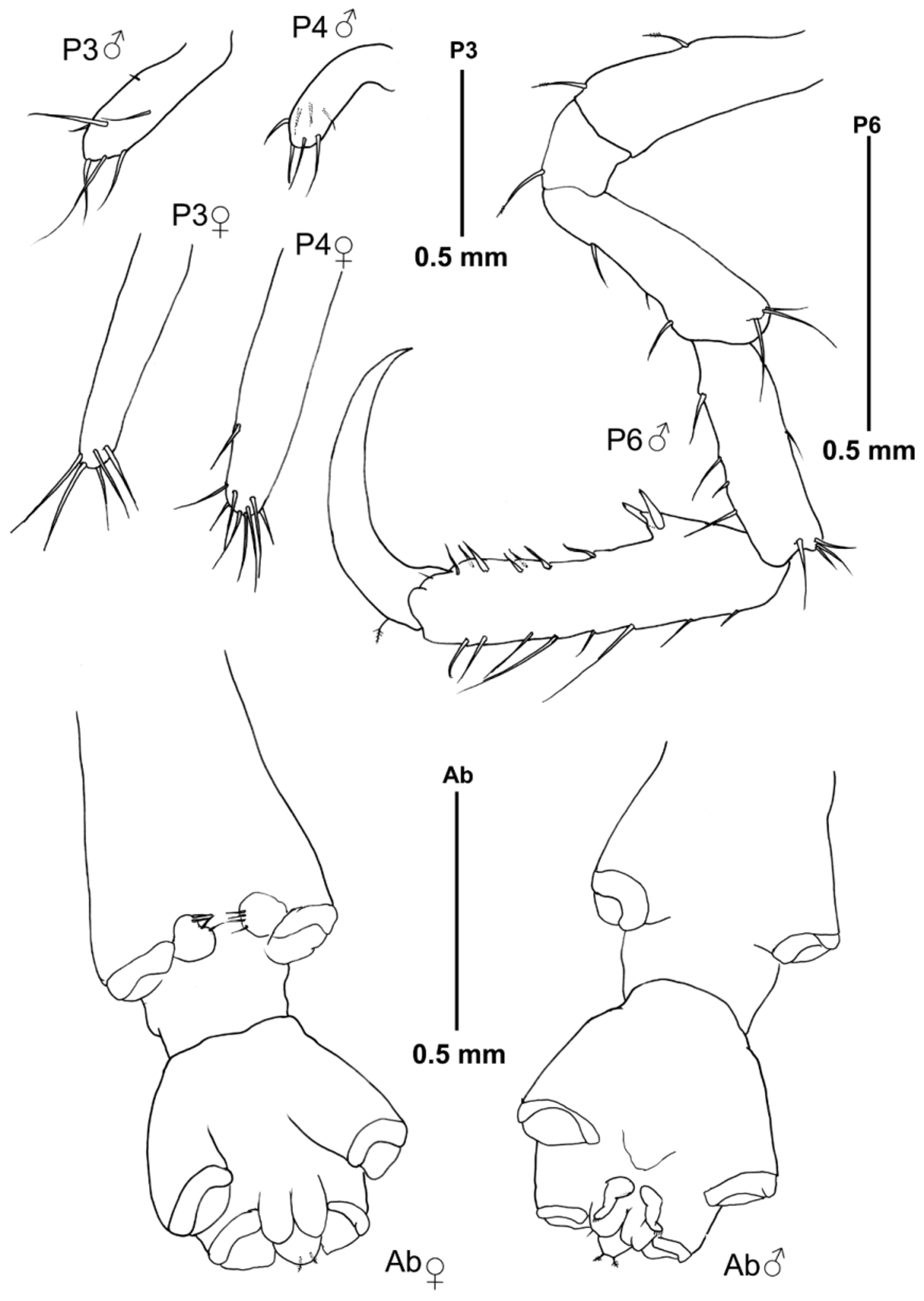
Pereopods 3–4 uniarticulate, about 0.25 x gill length Pereopods 5 and 7 missing; pereopod 6 7-articulate, robust, basis subequal in length than merus and carpus; propodus provided with a pair of proximal grasping spines, dactylus falcate.
Abdomen with a pair of 1-articulate appendages provided with fine setulae distally, pair of lobes and single dorsal lobe with pair of plumose setae.
Female (MNHN Am -5253).
Body length 5.8 mm. 2 dorsal projections medially on the head and pereonites 2 and 3, together with an additional dorsal projection distally on pereonites 2 and 3 and two medial protuberances on pereonite 4. A small projection antero laterally on pereonite 2, together with an acute projection (larger than in male) near the coxa of gnathopod 2 ( Fig. 4 View FIGURE 4 B). Antenna 1 flagellum 10-articulate. Gills less elongate than in male, and pereopods 3 and 4 slightly larger than in male. Palm of gnathopod 2 straight, with a small proximal projection provided with a grasping spine. Oostegites on pereonite 3 and 4 with inner margin setose. Abdomen lacking appendages, with pair of lateral lobes and single dorsal lobe with two plumose setae.
Remarks (see also remarks under Pseudoprotella phasma ). The only information available of M. africana correspond with the original description of Mayer (1903) based on a male and female collected from Djibouti, Somaliland, and a record of the species in the Gulf of Aqaba provided by Ruffo (1959). Mayer (1903) only provided the lateral view figure of the male and the female and his description is very concise. During a visit of one of the authors of the present study (JMGG) to the Muséum nacional d’Histoire naturelle, Paris, the type material of this species was located. Both the male and the female were in agreement with the lateral view figures provided by Mayer, and detailed figures and description are provided in the present study.
The generic diagnosis of Metaprotella was recently defined by Takeuchi & Lowry (2007) based on the description of newly collected material of Metaprotella haswelliana (Mayer, 1882) , the type species of Metaprotella . Lim & Takeuchi (2012) revised the diagnosis of Metaprotella in order to include variation in the suture between head and pereonite 1 and reported the following diagnostic characters for the genus: (1) Head fused (suture present or vestigial as slight concaved area) with pereonite 1; (2) Antenna 1 well developed, flagellum with more than 2 articles; (3) Antenna 2 well developed, flagellum with 2 articles; (4) Mandible well developed, molar present, well developed, palp 3-articulate with setal formula 1-x -y -1 or 1-x -1; (4) Maxilliped well developed, inner plate smaller than outer plate, outer plate well developed, palp article 3 with distal projection, palp article 4 well developed; (5) Pereonites 6 and 7 completely fused (dorsal suture absent); (6) Pereopods 3 and 4 vestigial, with 1 article; (7) Pereopod 5 well developed, with 7 articles, with sparse, short setae and well -developed dactylus; (8) Pereopods 6 and 7 well developed, with 7 articles; (9) Gills on pereonites 3 and 4; (10) Pleopods absent; (11) Uropods 2 pairs, biarticulate, unirramous and vestigial; (12) Telson (dorsal lobe) present.
Metaprotella africana agree with all the generic diagnostic characteristics except for the projection of the third article of the maxilliped palp, which is lacking in this species. In this sense, the diagnosis should be modified since the distal projection can be present or not depending on the species. Similarly, Takeuchi & Lowry (2007) and Lim & Takeuchi (2012) also considered in the diagnosis pereopods 3 and 4 uniarticulate. This is valid for all the species, including M. africana , except for M. tanzaniensis Guerra-Garcia, 2002b , which seem to have 2-articulate pereopods 3 and 4 (Guerra-Garcia, 2002b: p. 914, fig. 4A, B)
The diagnosis of the abdomen in the genus is not clear, since the second pair of uropods is difficult to assign in Metaprotella species. Even in M. haswelliana , the type species of the genus, the second pair of vestigial uropods reported in the diagnosis and description is not distinguishable in the illustrations (see Takeuchi & Lowry, 2007: p. 15, fig. 3h). In descriptions of other Metaprotella species, this second pair is also not recognizable, as shown, e.g, in McCain, 1968 (p.80, fig. 39i,j), Guerra-Garcia, 2002b (p. 914, fig. 4d), Guerra-Garcia, 2003b (p. 11, fig. 7f), Lim & Takeuchi, 2012 (p. 28, fig. 3) and Momtazy & Sari, 2013 (p. 199, fig. 2). Furthermore, the first pair of uropods is not clearly biarticulated in M. haswelliana , and is indeed clearly uniarticulate in other species of Metaprotella (see e.g. Müller, 1990: p. 840, fig. 57; Laubitz, 1991: p. 114, fig. 10G; Guerra-Garcia, 2002b: p.914, fig. 4d; Lim & Takeuchi, 2012: p. 28, fig. 3; Momtazy & Sari, 2013: p. 199, fig. 2). Consequently, a further detailed study of the abdomen in Metaprotella species should be conducted to properly clarify the diagnostic characters of abdominal appendages. On the other hand, the male penes are not clearly distinguishable in several species of Metaprotella . Although they can be recognized in M. sandalensis (Lim & Takeuchi, 2012: p. 28, fig. 3), they are not described for M. haswelliana (Takeuchi & lowry, 2007). The redescription of the present study show that the abdomen in M. africana is also complicate, the pair of appendages (uropods) is clearly uniarticulate, but the penes are not clearly differentiated in the male specimen.
Larsen (1997) and Guerra-Garcia (2003b) provided keys to the world species of Metaprotella . The genus presently comprises 11 species: M. haswelliana (Mayer, 1882) the type species of the genus, redescribed in detail based on material from Western Australia by Takeuchi & Lowry (2007), M. excentrica Mayer, 1890 described from Pamban Bridge, India and recently found also by Guerra-Garcia et al. (2010), M. problematica Mayer, 1890 described also from Pamban Bridge, India, M. sandalensis Mayer, 1898 , recently redescribed based on newly collected specimens from Lifou Island, New Caledonia, the type locality (Lim & Takeuchi, 2012), M. africana Mayer, 1903 redescribed in the present paper based on they type material designated by Mayer from Djibouti, Somaliland, M. makrodactylos Stebbing, 1910 described from South Africa, M. hummelincki McCain, 1968 from Puerto Rico (fully illustrated by McCain, 1968), M. unguja Larsen, 1997 and M. tanzaniensis Guerra-Garcia, 2002b , both described from Tanzania, M. mauritiensis Guerra-Garcia, 2003b , from Mauritius and M. macoranicus ( Momtazi & Sari, 2013) described from the Persian Gulf and the Gulf of Oman.
Most of the Metaprotella species are fully illustrated and properly described, except for M. excentrica , M. problematica and M. makrodactilos , which need to be redescribed in detail. All of the species show pereonites 6 and 7 totally fused, agreeing with the main diagnostic character for the genus. Metaprotella africana can be distinguished from the remaining species of Metaprotella mainly by the arrangement of dorsal projections; the male is provided with a pair of dorsal acute projections on pereonite 2, and the female with 2 dorsal projections medially on the head and pereonites 2 and 3, together with an additional dorsal projection distally on pereonites 2 and 3 and two medial protuberances on pereonite 4. Unfortunately, only two specimens constitute the type material of Mayer, and the different pattern of body projections arrangement in male and female cannot be confirmed in other specimens as a constant character. Larsen (1997) also found differences in body projections between male and female of M. unguja . M. africana seems to be closer to M. tanzaniensis ; however, the present redescription of M. africana shows that male has no paired dorsal projections on head, while female has distinct dorsal projection. To the contrary to M. africana , in M. tanzaniensis , male has distinct dorsal projection on head, while female has no paired dorsal projections on head. So male of M. africana looks closer to female of M. tanzaniensis , and female of M. africana looks closer to male of M. tanzaniensis .
Metaprotella species have, so far, a restricted distribution, apart from Metaprotella sandalensis which is commonly distributed throughout the tropical West Pacific Coasts (Lim & Takeuchi, 2012). McCain & Steinberg (1970) provided a detailed list of all the forms (or varieties) described for M. sandalensis . Guerra-Garcia (2003b) pointed out that the different morphology attributed to Metaprotella sandalensis sensu lato (globally distributed along the Indo-Pacific) could reflect a complex of species and suggested that a molecular approach would probably reveal the existence of cryptic species inside a complex. Lim & Takeuchi (2012) redescribed M. sandalensis from the type locality in detail and provided the distinctive morphological characters for the species. Momtazi & Sari (2013) recently described M. macoranicus as a new member of this M. sandalensis complex. The existence of cryptic species in caprellids has been recently reported for the C. penantis complex based on molecular evidence (Cabezas et al., 2013). However, molecular studies have also revealed that some species, such as Caprella scaura sensu stricto, which show intraspecific variations in the dorsal tubercles, can truly show a global distribution and that the morphological variation is intraspecific ( Ros et al., 2014; Cabezas et al., 2014). Existence of a clear intrapecific variation in some caprellid species has been also supported by experimental evidence ( Aoki & Kikuchi, 1990). Therefore, detailed morphological studies examining numerous specimens to explore intraspecific variation, together with the use of molecular tools, are necessary to dilucidate if a species is really widely distributed (naturally or human mediated), or if represents a complex of different species.
Distribution. Type locality: Djibouti, Somaliland. Other record: Gulf of Aqaba


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Metaprotella africana Mayer, 1903
| Zeina, Amr F. & Guerra, José M. 2016 |
Metaprotella africana
| Ruffo 1959: 23 |
| Mayer 1903: 42 |