
Massinium toyoshiomaruae Yamana, Ogawa & Ohtsuka, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5023.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:E759FD85-1904-45A7-9A4D-56815FB9649B |
|
persistent identifier |
https://treatment.plazi.org/id/AD40CD45-4132-FFD8-FF02-FC71FB5DFCDD |
|
treatment provided by |
Plazi |
|
scientific name |
Massinium toyoshiomaruae Yamana, Ogawa & Ohtsuka |
| status |
sp. nov. |
Massinium toyoshiomaruae Yamana, Ogawa & Ohtsuka View in CoL sp. nov.
[New Japanese name: Toyoshio-hana-namako]
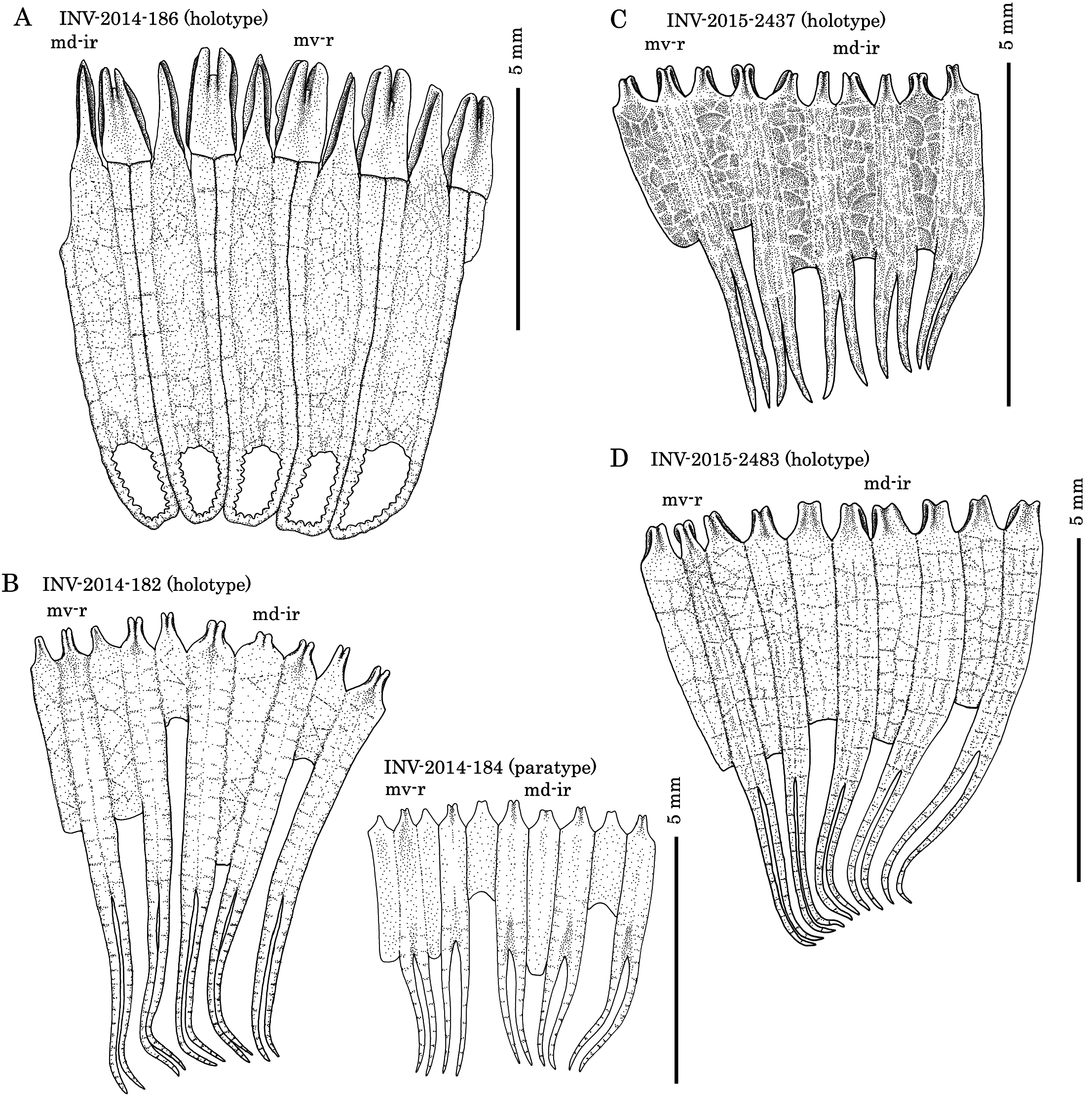
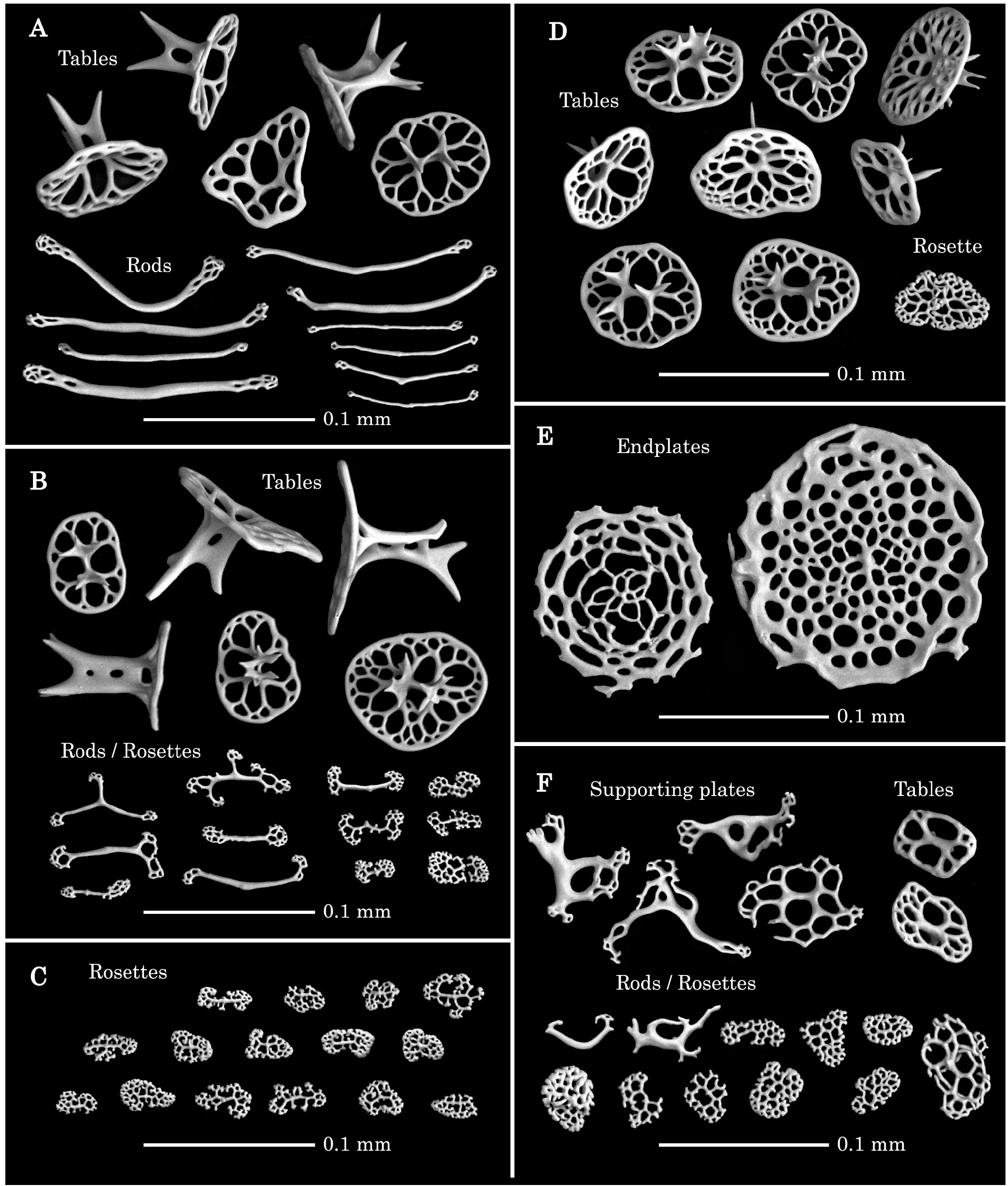
( Figs 2A View FIGURE 2 , 3A View FIGURE 3 , 4A–F View FIGURE 4 )
Material examined. Holotype, WMNH-INV-2014-186 (ST- 5 in 2014, anesthetized length 33 mm, width 10 mm, Fig. 2 View FIGURE 2 A-a). Paratypes: WMNH-INV-2015-2497 (ST- 9 in 2015, anesthetized, length 17 mm, width 6 mm, Fig. 2 View FIGURE 2 A-b); WMNH-INV-2015-2503 (ST- 10 in 2015, posterior half only, length 5 mm, width 3 mm, Fig. 2 View FIGURE 2 A-c); NSMT E- 12099-1 (ST- 10 in 2015, length 14 mm, width 5 mm, INSD accession number LC528687 View Materials , 514 bps, Fig. 2 View FIGURE 2 A-d) .
Diagnosis. Twenty tentacles arranged in double circles (10 + 10), comprising five pairs of large tentacles in the outer circle and five pairs of small tentacles in the inner circle. Calcareous ring elongate, tubular, with both radial and interradial plates, fragmented into a mosaic of small pieces and prolonged to the posterior, with posterior prolongations distally linked to others forming a loop. Bodywall lacking ossicles. Pedicels only with circular endplate, without supporting plates and rods.
Description. Body small, up to 33 mm, fusiform, curved, with both ends tapered and turned slightly upwards ( Fig. 2A View FIGURE 2 ); bodywall thick, soft. Living body color yellowish-brown with dark brown/gray pigmentation, preserved color paler than living color. Twenty tentacles arranged in double circles (10 + 10), comprising five pairs of large tentacles in the outer circle and five pairs of small tentacles in the inner circle. Tentacle bases and first bifurcations dark brown; tentacle trunks and tips transparent. One row of thin, short pharyngeal villi surrounding the oral opening. Oral disk brown. Retractable pedicels covering entire body at constant density, but forming two longitudinal rows along each radius in introvert region. Color of pedicels same or slightly paler than adjacent body. Pedicels becoming gradually smaller anterior and posterior from middle portion of body. Where they are denser. Ten anal papillae and five anal teeth on radii.
Calcareous ring elongate, tubular, with both radial and interradial plates fragmented into a mosaic of small pieces and prolonged posteriorly, with the posterior prolongations distally linked to prolongations of neighboring plates ( Fig. 3A View FIGURE 3 ). Polian vesicles paired, both in medio-ventral position, stone canal single, in mediodorsal. Well-matured gonad situated in mid-body, in two clumps, one on each side of the dorsal mesentery, most tubules branched. Gonad lacking ossicles.
Bodywall lacking ossicles. Tentacle ossicles comprising thin rods in tentacle branches, variously sized, somewhat curved, most with small numbers of distal perforations ( Fig. 4A View FIGURE 4 , Table 3). Tentacle bases and adjacent area of peri-oral and introvert skin with regular tables with oval discs possessing four large central and numerous small peripheral perforations, and two-pillared low spires mostly with one crossbeam and 2–6 apical spines ( Fig. 4A, B, D View FIGURE 4 , Table 3). Peri-oral skin ossicles comprising mostly thin-framed, irregular-shaped rods/rosettes, also regular tables in parts adjoining tentacle bases, disc shape of tables in tentacle and peri-oral skin resemble each other, however, mostly have spires higher than those of tentacles and with 2–3 crossbeams, ( Fig. 4B View FIGURE 4 , Table 3). Pharyngeal villi have many thin-framed rosettes ( Fig. 4C View FIGURE 4 , Table 3). Introvert possessing tables with the same disc shape of those in tentacles and peri-oral area, however, most have low spires with one or no crossbeams ( Fig. 4D View FIGURE 4 , Table 3). We have several paratypes including smaller individuals, however, we cannot find the evidence supporting a hypothesis the specimens lose/modify their bodywall deposits with age: any individual with nothing of bodywall deposits.
Pedicels only with circular endplate, without supporting plates/rods, any kinds of supporting rods could not be observed even in the tips of the pedicels ( Fig. 4E View FIGURE 4 , Table 3). Anal papillae mostly with supporting plates in distal part, and also with small tables with low spires, and rosettes in basal part ( Fig. 4F View FIGURE 4 , Table 3). Skin around anus occasionally possessing regular tables.
Remarks. This species is characterized by the peculiar shape of calcareous ring and possessing 20dendriform tentacles. Presently, these features are the principal features of the genus Massinium and Neotyonidium, former genus is well-defined from the latter by the lacking of bodywall table ossicles and includes 12 nominal species (WoRMS 2021a) (Table 7). In Japanese waters only two species of this genus, M. magnum ( Ludwig, 1882) , M.japonicum ( Yamana, Ota & Kohtsuka, 2020) has been reported ( Nomura 2004; Yamana, Ota & Kohtsuka, 2020). The new species M. toyoshiomaruae lacks bodywall ossicles and pedicel supporting plates/rods, and these features readily distinguish it from all congeners (Table 7). Among the 12 Massinium species , the following nine possess bodywall ossicles: M. albicans Samyn, Thandar & Vandenspiegel, 2010 having rosettes and pseudobuttons, M. arthroprocessum ( Thandar, 1989) with flattened rods, M. bonapartum O’Loughlin in O’Loughlin et al. (2014) with scales and tables, M. dissimilis ( Cherbonnier, 1988) with tables and pseudobuttons, M. granulosum Samyn, Thandar, & Vandenspiegel, 2010 bearing granuliform rods and pseudobuttons, M. japonicum ( Yamana, Ota & Kohtsuka, 2020) with rosettes ( Yamana, Ota & Kohtsuka, 2020), M. keesingi O’Loughlin in O’Loughlin et al. (2014) with tables, M. maculosum Samyn & Thandar, 2003 with rosettes, and M. magnum ( Ludwig, 1882) with rosettes and pseudobuttons. However, three Australian species, M. melanieae O’Loughlin in O’Loughlin et al. (2012), M. vimsi O’Loughlin in O’Loughlin et al. (2012), and M. watsonae O’Loughlin in O’Loughlin et al. (2012), are frequently lacking mid-bodywall ossicles. Although the new species is also lacking mid-bodywall ossicles, it differs from these three species by the absence of the pedicel supporting rods and the presence of the supporting rods in the tips of the pedicels ( O’Loughlin et al, 2012).
Distribution. In the Seto Inland Sea and western part of the Sea of Japan, depth 12– 73 m. The Bisan-Seto Strait, eastern part of the Seto Inland Sea, sand bottom, depth 12–16 m (ST- 5 in 2014); western part of the Sea of Japan, sand bottom, depth 72–74 m (ST-9, 10 in 2015).
Etymology. The specific name toyoshiomaruae alludes to the name (TR/V) TOYOSHIO MARU of Hiroshima University.
Molecular data comparison. In BLAST searches, the three closest hits obtained for the partial COI gene sequence were those of Neothyonidium sp. (INSD accession number KF142179 View Materials and KF142176 View Materials , 86.5% and 86.2% respectively) and Massinium magnum ( KF142162 View Materials , 84.6%). The results of BLAST searches confirmed that the present species is genetically related to M. magnum , and supported that the present species belongs to the genus Massinium and it and is distinct from M. magnum . The results of BLAST searches also showed that the present species is similar to Neothyonidium sp. However, the detailed morphologies of voucher specimens of Neothyonidium sp. ( KF142179 View Materials and KF142176 View Materials ) are unavailable, and INSD sometimes includes misidentification; thus, morphological studies to confirm the relatedness of these species are required. However, this result is quite acceptable because the previous nominated species of Massinium were put in Neothyonidium until Samyn and Thandar (2003) erected Massinium .

| NSMT |
National Science Museum (Natural History) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.