
Mantidactylus marintsoai, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504399 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FF55-FF42-4F13-4F4F4C437B2E |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus marintsoai |
| status |
sp. nov. |
Mantidactylus marintsoai sp. nov.
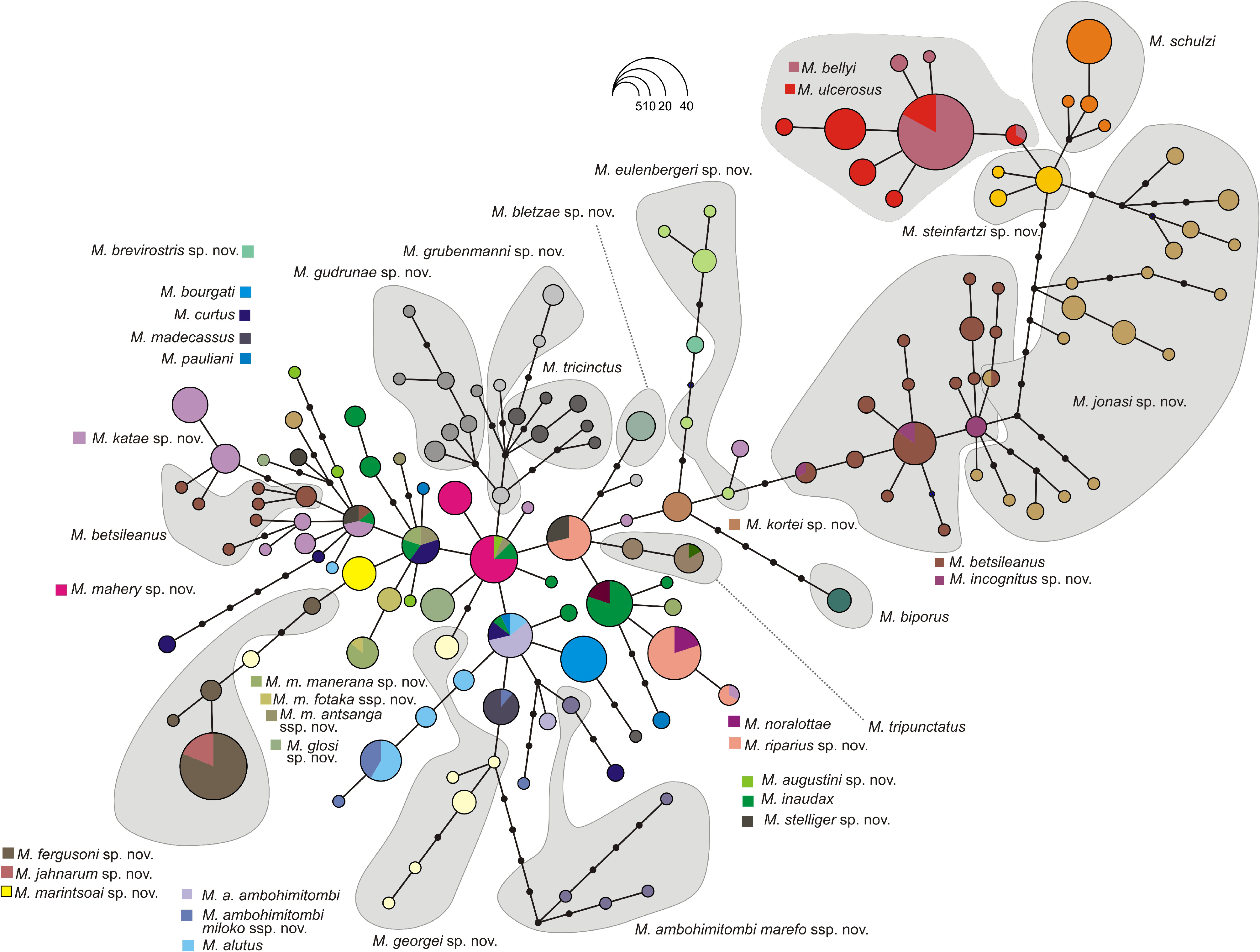
Identity and justification.—This lineage was newly identified in this study. It consists of frogs from the North East of Madagascar that form a mitochondrially homogeneous lineage that probably is the sister group of M. georgei ( Fig. 2 View FIGURE 2 ; M. marintsoai sp. nov. is not included in the phylogenomic tree), with which it shares a unique Rag-1 haplotype ( Fig. 4 View FIGURE 4 ). Despite the lack of unambiguous morphological differences to M. georgei and the absence of bioacoustic data for M. marintsoai sp. nov., we here name this lineage as new species, given its enormous genetic divergence (7.6–9.3% uncorrected 16S distance to M. georgei ).
Holotype.— CURSA-A 0033/2021 (field number THC301), adult male, collected by T.R. Fulgence, D.A. Martin, R. Randriamanantena, R. Botra, E. Befidimanana, A. Wurz, K. Osen, H. Kreft, A. Andrianarimisa, and F. M. Ratsoavina on 30 August 2018 in the eastern part of Marojejy National Park ‘Bangoabe’ (14.4467°S, 049.8251°E, ca 225 m a.s.l.), Sava Region, Madagascar. GoogleMaps A 16S barcode sequence of the holotype was obtained in this study and was included in the analysis.
Paratypes.—A total of six paratypes: CURSA-A 0034/2021 (THC248, THC245, THC246), three adult females, collected by T.R. Fulgence, D.A. Martin, R. Randriamanantena, R. Botra, E. Befidimanana, A. Wurz, K. Osen, H. Kreft, A. Andrianarimisa, and F. M. Ratsoavina on 18 January 2018 in a stream of Fokontany Andramanolotra/Sambava (13.9953°S, 050.0711°E, ca 54 m a.s.l.); GoogleMaps CURSA-A 0036/2021 (THC354), adult female, and THC355, juvenile, collected by the same team on 22 November 2018 in the same location; GoogleMaps CURSA –A 0035/2021 (THC233) adult female, collected by T.R. Fulgence, D.A. Martin, R. Randriamanantena, R. Botra, E. Befidimanana, A. Wurz, K. Osen, H. Kreft, A. Andrianarimisa, and F. M. Ratsoavina on 3 February 2018 in a stream in the Antsirabe-Nord / Vohemar, Fokontany of Bemanevika (13.9864°S, 049.9519°E, ca 61 m a.s.l.) GoogleMaps .
Diagnosis.— Mantidacytlus marintsoai sp. nov. is a member of the M. fergusoni clade, and is probably the sister lineage of M. georgei , according to the 16S tree (it is not included in the phylogenomic analysis). See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of a moderate to large body size (male SVL 29 mm, female SVL 35–39 mm), moderately tubercular dorsal skin, relatively large tympanum size in males (13% of SVL), absence of white spots on flanks, and absence of a white marking on the snout tip, distinguishes M. marintsoai sp. nov. from most species of the other clades ( Table 4 View TABLE 4 ). Two species from the M. ulcerosus clade ( M. ulcerosus and M. bellyi ) can be morphologically similar; these two species occur in the Sambirano and North West regions, and the distribution area especially of M. bellyi may overlap with that of M. marintsoai sp. nov. which is known from the North East; a distinction may be possible by the apparently more strongly developed foot webbing in M. bellyi and M. ulcerosus ( Table 4 View TABLE 4 ). Mantidactylus marintsoai sp. nov. shows similarities to species of the M. betsileanus clade but does not appear to occur sympatrically with any of them; in general, it has a more tubercular dorsum and seems to lack a distinct white marking on the snout tip which characterizes many species of the M. betsileanus clade ( Table 4 View TABLE 4 ). Within the M. fergusoni clade, a bioacoustic comparison is not possible due to the lack of call data for M. marintsoai sp. nov.; the species might be distinguished from M. fergusoni , M. georgei and M. jahnarum by less developed foot webbing ( Table 4 View TABLE 4 ), and seems to differ from many specimens of M. georgei and M. jahnarum by the lack of a clear white marking on snout tip. For a distinction from other new species described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. marintsoai sp. nov. in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Description of the holotype.— Adult male in good state of preservation ( Fig. 48 View FIGURE 48 ). For measurements, see Table 8. Body rather stout. Head longer than wide. Snout rather pointed. Nostrils directed laterally, slightly protuberant, nearer to tip of snout than to eye. Canthus rostralis distinct, loreal region slightly concave. Tympanum distinct, large, round, horizontal diameter of tympanum 97% of horizontal eye diameter. Supratympanic fold distinct, beginning straight above, with rather distinct bend midway towards insertion of forelimb. Tongue ovoid, distinctly bifid posteriorly.Vomerine teeth form two elongate aggregations, positioned posterolateral to choanae. Choanae rounded.
Subarticular tubercles single. Outer metacarpal tubercle not recognisable, inner metacarpal tubercle present. Fingers without webbing. Relative length of fingers: I<II<IV<III.
Finger discs slightly enlarged. Nuptial pads absent. Foot slightly shorter than tibia (98%). Lateral metatarsalia separated. Inner metatarsal tubercle present. Outer metatarsal tubercle not recognisable. Webbing formula: 1(1), 2i(1), 2e(0.5), 3i(2), 3e(1), 4i(2), 4e(2), 5(1). Relative length of toes: I<II<V<III<IV. Skin on the upper surface smooth, dorsolaterally smooth. Ventral side smooth. Femoral glands small and distinct in external view.
Colour in preservative: dorsally brown, with two distinct light spots. Forelimbs light brown with 2–3 dark brown crossbands on hand and arm. Hindlimbs light brown with distinct dark brown crossbands. Inguinal region with few scattered whitish spots. Snout tip with a light spot (width 1.1 mm, height 1.2 mm), tympanum region slightly lighter than remaining head sides. Venter beige with light brown mottling, throat darker than belly with brown mottling. Lower lip with distinct alternating light and brown spots. Toe discs dark brown.
Variation.—Variation in measurements is given in Table 8. See Fig. 55 View FIGURE 55 for colouration in life and its variation. Some individuals have mid-dorsal stripes ( Fig. 55b View FIGURE 55 ). There may be pronounced sexual size dimorphism, but sample sizes are small (confirmed male SVL 29.0 mm [n = 1] vs confirmed female SVL 34.6–38.7 mm [n = 5]). Males seem to have a larger tympanum than females (HTD/ED ratio is 103% in the only known male, 50–71% in females). Femoral glands in life, in the only known male, are relatively small and indistinct, and not conspicuously coloured; tiny rudimentary glands are recognisable in females.
Natural history.—Found in rainforest patches of the North East of Madagascar. All individuals were found in or near streams in rainforest, including degraded primary forest, secondary forest, and narrow riparian forest fragments. In 2019, the riparian forest beside the village of Bemanevika was felled.
Call.—The call of this species has not been recorded.
Tadpoles.—The tadpole of this species has not been described.
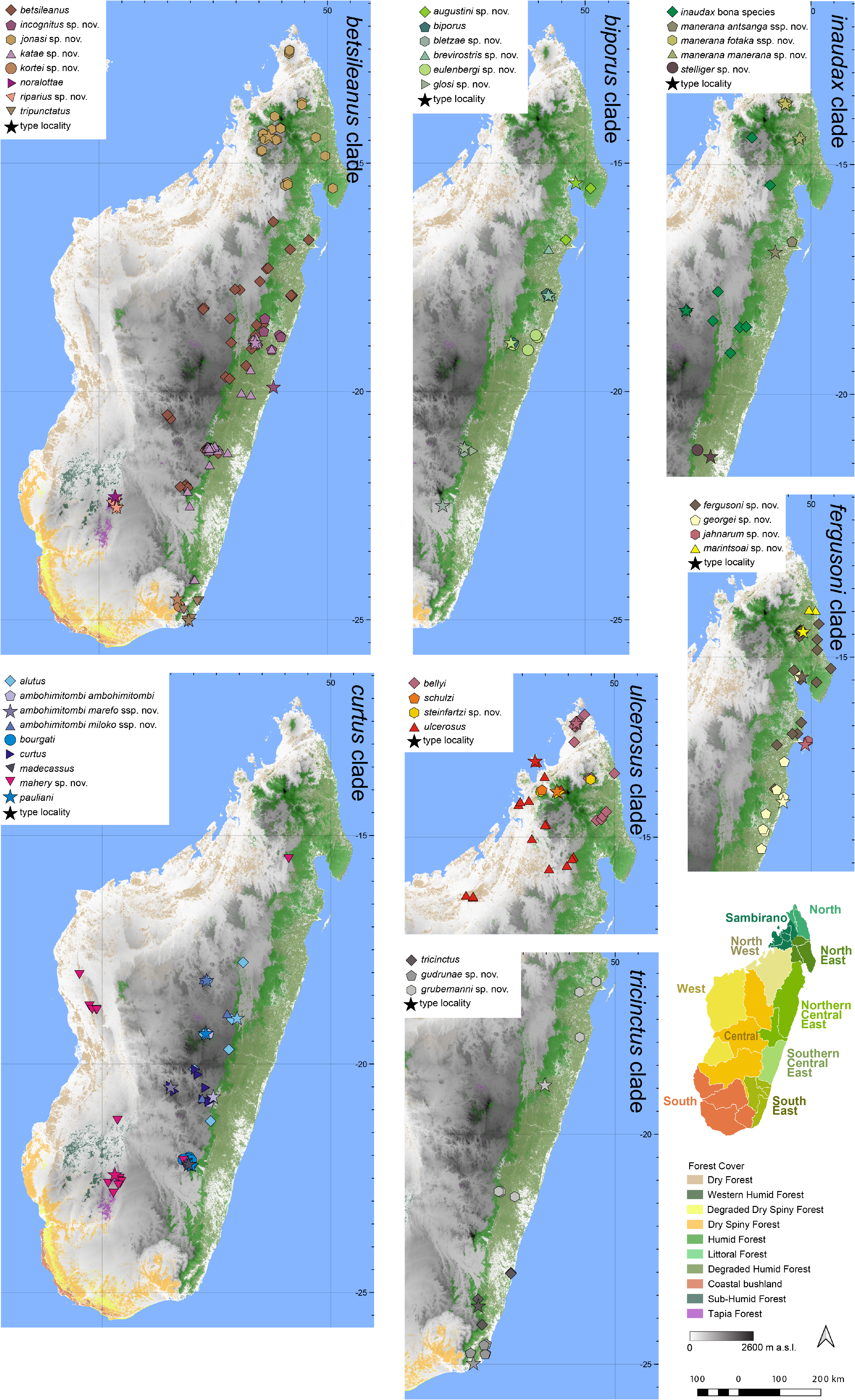
Distribution.— Apparently endemic to a rather small area of the North East of Madagascar ( Fig. 7 View FIGURE 7 ).
This species is known from Marojejy, Andramanolotra, and Bemanevika village (a Fokontany of AntsirabeNord, in the Vohemar district). Elevation range: 16–273 m a.s.l.
Etymology.—T.R. Fulgence wishes to dedicate this species to his father, the late Marintsoa, in recognition of his support of his son’s scientific career and pursuit of a PhD degree. The name is used as a noun in apposition.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |