
Mantidactylus mahery, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504356 |
|
persistent identifier |
https://treatment.plazi.org/id/C2BCF600-E15B-41E6-A27C-3CD8F3B6B040 |
|
taxon LSID |
lsid:zoobank.org:act:C2BCF600-E15B-41E6-A27C-3CD8F3B6B040 |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus mahery |
| status |
sp. nov. |
Mantidactylus mahery sp. nov.
Identity and justification.—This lineage has been considered as confirmed candidate species M. sp. 14 by Vieites et al. (2009) and M. sp. Ca14 by Perl et al. (2014).
This is a relatively large-sized species of Brygoomantis fromwesternMadagascarwithamorphologysuperficially similar to M. ulcerosus which, however, is not its closest relative and concordantly differs in mitochondrial and nuclear genes and in advertisement call structure.
Both species occur syntopically at least at one site (Makira). Although M. mahery sp. nov. mainly occurs in western Madagascar unlike the other species of the M. curtus clade which live in the central highlands, the phylogenomic data unambiguously support its inclusion in the group. Even more surprising, the phylogenomic tree places the species sister to M. pauliani , which is a montane endemic from the Ankaratra Massif that differs in numerous morphological characters. The status of M. mahery sp. nov. as a separate species is well supported by multiple lines of evidence.
Holotype. — ZSM 23/2004 (field number FGZC 37), adult male collected by F. Glaw, M. Puente, R. Randrianiaina, and M. Teschke (née Thomas) on 21 January 2004 in Isalo at a creek near Ranohira (22.5856°S, 045.3997°E, 813 m a.s.l.), Ihorombe Region, Madagascar.
A 16S barcode sequence of the holotype is available from GenBank (accession AY848286 View Materials ).
GoogleMapsParatypes.—A total of 12 paratypes GoogleMaps : ZSM 25/2004 ( FGZC 39 ), adult male, and ZSM 26/2004 ( FGZC 42 ), adult female, with the same collection data as the holotype; ZSM 567/2009 ( ZCMV 11457 ) and ZSM 569/2009 ( ZCMV 11484 ), two adult females, collected by M. Vences, D.R. Vieites, F.M. Ratsoavina, R.D. Randrianiaina, E. Rajeriarison, T. Rajofiarison, and J. Patton on 20 June 2009 in Sahaovy (‘Camp 0’), Makira (15.4889°S, 049.0785°E, 607 m a.s.l.) GoogleMaps ; ZSM 9/2006 ( FGZC 682 ), adult male, and ZSM 61/2006 ( FGZC 793 ), adult female, collected by F. Glaw, J. K̂hler, P. Bora, and H. Enting on 18 and 23 March 2006, respectively, at Antranopasazy (‘Camp 1’), Tsingy de Bemaraha National Park (18.7086°S, 044.7189°E, 146 m a.s.l.) GoogleMaps ; ZSM 134/2006 ( FGZC 938 ), ZSM 135/2006 ( FGZC 939 ), two adult males, and ZSM 136/2006 ( FGZC 940 ), adult female, collected by F. Glaw, J. K̂hler, P. Bora, and H. Enting on 31 March 2006 at Andafiabe on the Beboka River (‘Camp 2’), Tsingy de Bemaraha National Park (18.7842°S, 044.7794°E, 177 m a.s.l.) GoogleMaps ; ZSM 927/2003 ( FG / MV 2002.1421 ), putative male, ZSM 941/2003 ( FG / MV 2002-1485 ), adult male, and ZSM 942/2003 ( FG / MV 2002-1486) , adult female, collected by G. Aprea, M. Puente, L. Raharivololoniaina, M. Teschke (née Thomas), and D.R. Vieites between 29 January and 1 February 2003 at Hotel Isalo Ranch (22.5929°S, 045.3928°E, ca 800 m a.s.l.) GoogleMaps .
Diagnosis.— Mantidactylus mahery sp. nov. is a member of the M. curtus clade as revealed by the phylogenomic analysis, and sister to the morphologically strongly different M. pauliani . While all other species in the M. curtus clade occur on the central plateau of Madagascar, M. mahery is distributed in western Madagascar, including some rather arid areas where it appears to be the only Brygoomantis present, and it is also present in one locality in the North West (western slope of Makira). See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of a large body size of up to 49 mm, slightly granular skin with (weakly expressed) dorsolateral ridges, absence of white spots on flanks and of white marking on snout tip, and short pulsed advertisement calls emitted in regular series distinguishes the new species from species of the other clades. In the North West it can occur syntopically with the similarly sized M. ulcerosus which however has a distinctly more tubercular dorsal skin, and more pulses per note in advertisement calls. Within the M. curtus clade, the new species differs by its larger tympanum diameter in males (11–13% of SVL) from all other species ( Table 4 View TABLE 4 ). Mantidactylus alutus , M. madecassus and M. pauliani furthermore are smaller and have a shorter snout ( Table 4 View TABLE 4 ). For detailed distinction from other new species described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. alutus in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Description of the holotype. —Adult male in good state of preservation ( Fig. 9 View FIGURE 9 ). Tongue excised as tissue sample (tongue no longer present); femoral glands partly detached for examination in internal view. Body rather slender (in this, differing from many other specimens of this species which are stouter). Head as wide as body. Snout rounded in dorsal and lateral views. Nostrils directed laterally, slightly protuberant. Nostrils nearer to tip of the snout than to eye. Canthus rostralis weak, slightly concave. Loreal region weakly concave. Tympanum distinct, large, elliptical, diameter about 93% of eye diameter. Supratympanic fold present, beginning straight,with a rather distinct bend midway towards forelimb insertion. Maxillary teeth present. Vomerine teeth present in two rounded aggregations, positioned posterolateral to choanae. Choanae rounded. Subarticalur tubercles single. Outer metacarpal tubercle present, inner metacarpal tubercle present. Fingers without webbing. Relative length of fingers: I=II<IV<III. Finger discs slightly enlarged. Nuptial pads absent. Foot longer than tibia (115%). Lateral metatarsalia separated. Inner metatarsal tubercle present. Outer metatarsal tubercle not present. Webbing formula: 1(0.25), 2i(1), 2e(0), 3i(1), 3e(0.5), 4i(2), 4e(1), 5(0.5). Relative length of toes: I<II<V<III<IV. Skin on the upper surface quite smooth with very few scattered granules on flanks. Ventral side smooth. Femoral glands present, in external view with a distal ulcerous macrogland and a large proximal granular gland field.
Colour in preservative: dorsum dark brown. Forelimbs brown with poorly defined darker markings. Hindlimbs brown with poorly defined darker crossbands. Inguinal region without whitish spots. Snout tip without a light dot. Venter beige with brown mottling, throat darker than belly. Lower lip with alternating light and brown spots.
Toe discs dark. Colour in life of holotype unknown.
Variation. —Variation in measurements is given in Table 5 View TABLE 5 . See Fig. 21 View FIGURE 21 for colouration in life and its variation.
In life, dorsum is brown with distinct darker markings.
Dark band between eyes is present. Forelimbs brown with very indistinct darker markings; hindlimbs with indistinct darker crossbands. Belly beige; throat with distinct white and brown mottling. A longitudinal white line on abdomen and throat is present. Femoral glands distinctly orange.
There is moderate sexual size dimorphism(confirmed male SVL 29.2–37.2mm [n =7] vs confirmed female SVL 34.2–48.6 mm [n = 6]). Horizontal tympanum diameter is 80–100% of eye diameter in males and 69–86% of eye diameter in females. Skin on the back with very few indistinct tubercles on the flanks. Colour on the back varies from light brown with distinct darker markings (e.g. ZSM 927/2003) to uniformly dark brown. Two dark spots on the back at the level of forelimb insertion always more or less distinctly present, except in the holotype (ZSM 23/2004), whose colour is too dark to see any markings.A dark brown more or less triangular band between eyes is always present. A light vertebral band or line is not present. An indistinct light dot on the snout tip is never present except in ZSM 25/2004. Lower lip with more (e.g. ZSM 23/2004) or less (e.g. ZSM 25/2004) distinct alternating light and brown spots. Venter and throat from uniformly beige with faint markings (e.g. ZSM 26/2004) to dark brown mottled (e.g. ZSM 23/2004). A longitudinal light median line on abdomen and throat is present in ZSM 25/2004. Hindlimbs always distinctly striped (e.g. ZSM 941/2003) except in ZSM 23/2004 where hindlimbs are striped indistinctly dark-brown. Forelimbs brown with irregular darker markings and stripes. Femoral glands of males large and prominent with a clear proximal granular gland field in ZSM 25/2004 and ZSM 941/2003, in ZSM 23/2004 and ZSM 927/2003 less prominent with indistinct proximal granular gland field. In external view a central depression in the middle of the femoral gland can be seen, thus indicating a distal ulcerous macrogland. In females femoral glands are always small but distinctly present (e.g. ZSM 942/2003), but a proximal granular gland field is never present. In life, males in reproductive state have femoral glands orange coloured ( Fig. 21b, h View FIGURE 21 ), and sometimes ( Fig. 21b View FIGURE 21 ) the proximal granular gland field is larger and more prominent than the distal ulcerous macrogland, which is uncommon among mantellines; the granular gland fields on the two opposite thighs contact each other medially.
Natural history.—The species is known from various sites in the West and North West of Madagascar, reaching into the South at Isalo. It has been found along running water (including very slowly running streams) in and outside of forest. At the western slope of the Makira Reserve it was found with M. ulcerosus and M. jonasi sp. nov. (see below) along a relatively large stream in degraded remnants of rainforest.
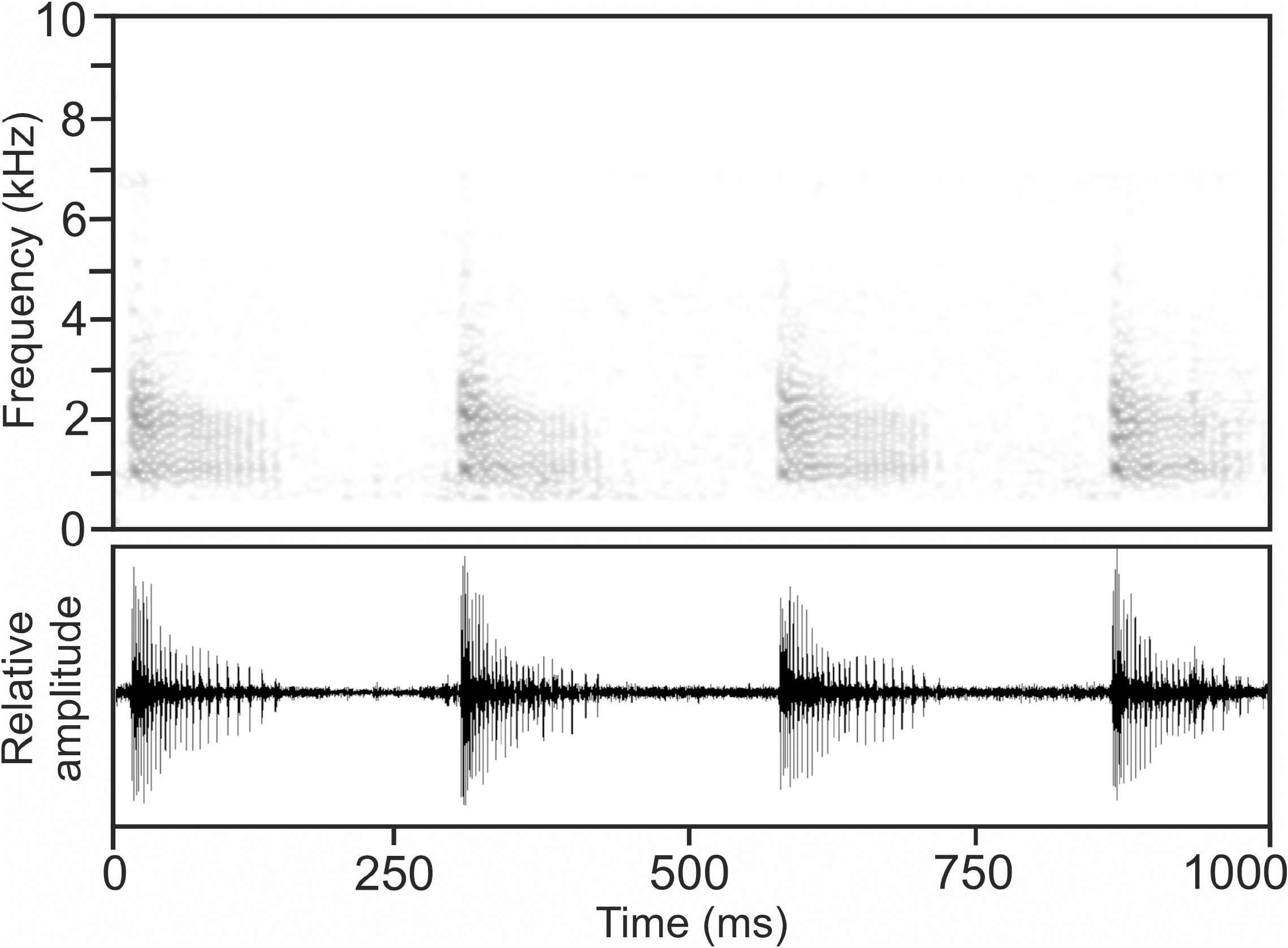
Calls.— The advertisement call of M. mahery , recorded on 28 January 1994 at Isalo National Park, near Ranohira, 23.4°C air temperature ( Vences et al. 2006: CD 2, track 76), consists of a short, pulsed note, emitted in regular series at fast succession ( Fig. 22 View FIGURE 22 ). Pulse repetition rate is distinctly higher at the beginning of calls and significantly reduces after approximately one quarter of the call’s duration. Amplitude modulation is present, with highest call energy occurring at the beginning of the call and continuously decreasing towards its end. Numerical parameters of eight analysed calls are as follows: call duration (= note duration) 120– 144 ms (131.8 ± 8.7 ms); 21–26 pulses per note (24.1 ± 2.3); pulse duration 1–2 ms (1.6 ± 0.5); pulse repetition rate within notes 107.8–667.0 pulses/s (331.4 ± 205.3); dominant frequency 1004–1270 Hz (1151 ± 95 Hz); prevalent bandwidth 800–4500 Hz; call repetition rate (= note repetition rate) within regular series ca 211–218 calls/min.
Tadpoles.— The tadpole of this species has not been described.
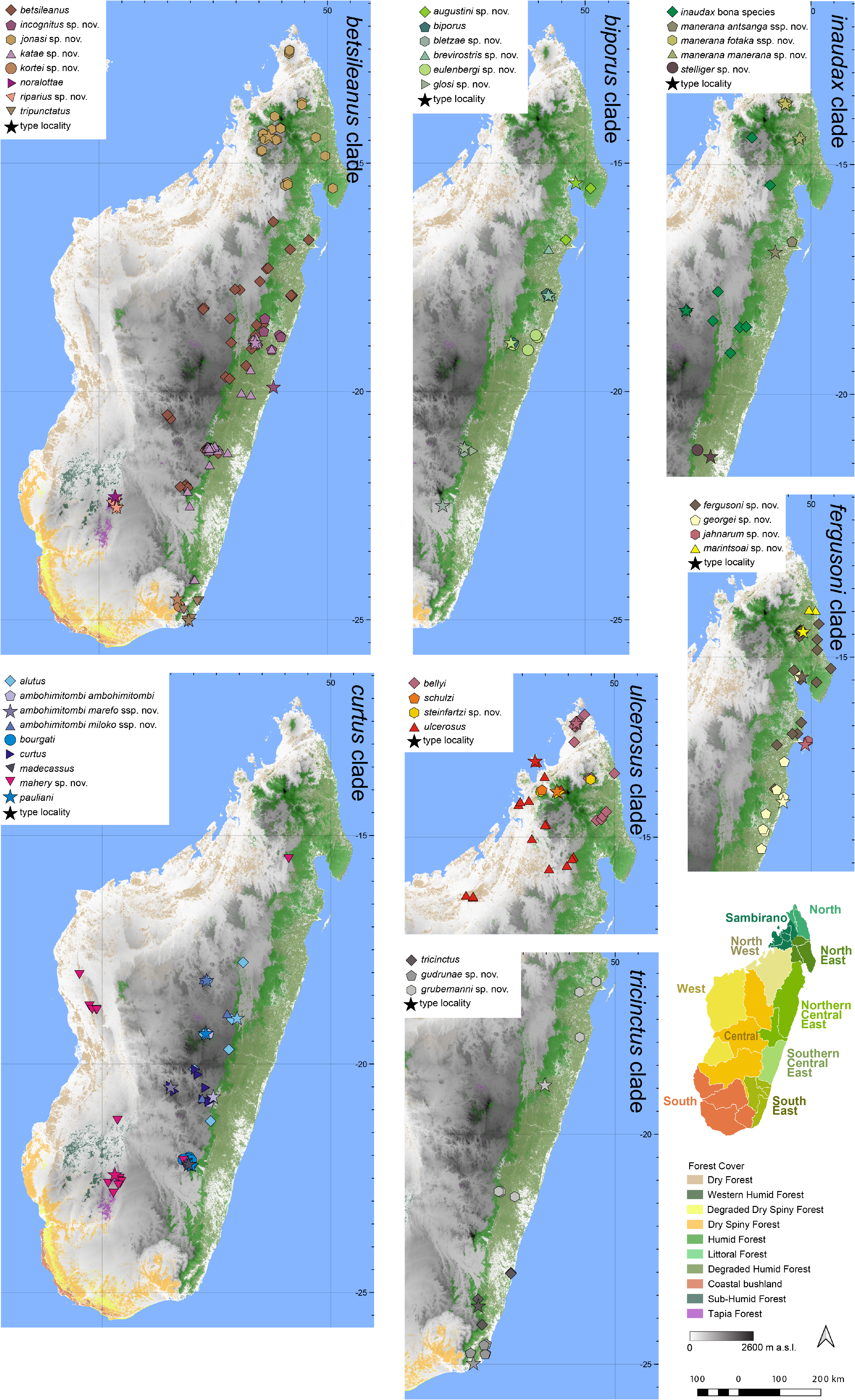
Distribution.— Endemic to an eclectic collection of disparate localities, mostly in the West of Madagascar, but reaching the eastern rainforest escarpment at Makira in the North West, and Tsaranoro in the Central region ( Fig. 7 View FIGURE 7 ). This species is known from Isalo (various localities), Forêt de Beanka, Makay, Makira West (Sahaovy, Camp 0), Tsaranoro, and Tsingy de Bemaraha (various localities). Elevation range: 120–960 m a.s.l.
Etymology. —The species name is derived from the Malagasy adjective mahery, meaning ‘big’ or ‘strong’, and refers to the rather stout body shape of this species. The name is used as a noun in apposition.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |