
Mantidactylus jonasi, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504383 |
|
persistent identifier |
https://treatment.plazi.org/id/5DEF04E9-81D7-4A71-A7F5-5EB1278711F4 |
|
taxon LSID |
lsid:zoobank.org:act:5DEF04E9-81D7-4A71-A7F5-5EB1278711F4 |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus jonasi |
| status |
sp. nov. |
Mantidactylus jonasi sp. nov.
Identity and justification.—This lineage previously was considered to represent a northern deeply divergent conspecific lineage of M. betsileanus , due to its superficial similarity in advertisement calls, and very similar morphology. However, closer investigation revealed that (i) the mitochondrial divergence is very high (5.1‒ 8.3%), (ii) there is almost no haplotype sharing with M. betsileanus in the nuclear gene Rag-1, (iii) the calls from various sites have a consistently lower pulse repetition rate and lower number of pulses per call, and (iv) the northern specimens are very slightly but consistently smaller than M. betsileanus . Taken together, this is reliable evidence for a distinctness at species level.
Holotype.— ZSM 86/2016 ( MSZC 0180 ), adult male, collected by M.D. Scherz, J. Borrell, L. Ball, T. Starnes, E. Razafimandimby, D.H. Nomenjanahary, and J. Rabearivony on 9 January 2016 in Ampotsidy (14.42832°S, 048.72129°E, 1227 m a.s.l.), Sofia Region, Madagascar. GoogleMaps A 16S barcode sequence of the holotype was obtained in this study and was included in the analysis.
Paratypes.—A total of eleven paratypes GoogleMaps : ZSM 133/2018 ( MSZC 0714 ) and ZSM 134/2018 ( MSZC 0715 ), two adult males, collected by M.D. Scherz, J.H. Razafindraibe, A. Razafimanantsoa, O. Randriamalala, S.M. Rasolonjavato, R. Tiavina, and A. Rakotoarison on 30 November 2017 at high elevation on Montagne d’Ambre GoogleMaps (12.58516°S, 049.14875°E, 1225 m a.s.l.); UADBA uncatalogued ( MSZC 0617 ), adult male, collected by M.D. Scherz, J.H. Razafindraibe, A. Razafimanantsoa, O. Randriamalala, S.M. Rasolonjavato, R. Tiavina, and A. Rakotoarison on 16 November 2017 at intermediate elevation on Montagne d’Ambre (12.52660°S, 049.16806°E, 1071 m a.s.l.) GoogleMaps ; UADBA uncatalogued ( MSZC 0729 ), unsexed individual, collected by M.D. Scherz, J.H. Razafindraibe, A. Razafimanantsoa, O. Randriamalala, S.M. Rasolonjavato, R. Tiavina, and A. Rakotoarison between 2–4 December 2017 at high elevation on Montagne d’Ambre (12.59247°S, 049.15302°E, 1372 m a.s.l.) GoogleMaps ; ZSM 2220/2007 ( FGZC 1368 ) and ZSM 2222/2007 ( FGZC 1372 ), two adult males, collected by F. Glaw, P. Bora, H. Enting, J. K̂hler, and A. Knoll on 11–12 March 2007 near the Gîte d’Étape on Montagne d’Ambre (ca 12.527°S, ca 049.172°E, ca 1040 m a.s.l.) GoogleMaps ; ZSM 1773/2010 ( ZCMV 12603 ) and ZSM 1774/2010 ( ZCMV 12604 ), two adult males, collected by M. Vences, D. Vieites, R.D. Randrianiaina, F.M. Ratsoavina, S. Rasamison, A. Rakotoarison, E. Rajeriarison, and T. Rajoafiarison on 29 June 2010 in a forest fragment between Bealanana and Antsohihy (14.72145°S, 048.56272°E, 1187 m a.s.l.) GoogleMaps ; ZSM 89/2016 ( MSZC 0194 ), adult male, collected by M.D. Scherz and M. Rakotondratisma on 14 January 2016 in a forest called Andranonafindra between Bealanana and Antsohihy (14.73600°S, 048.54831°E, 1180 m a.s.l.) GoogleMaps ; ZSM 551/2009 ( ZCMV 11462 ), adult male, collected by M. Vences, D.R. Vieites, F.M. Ratsoavina, R.D. Randrianiaina, E. Rajeriarison, T. Rajofiarison, and J. Patton on 20 June 2009 at Sahaovy (‘Camp 0’), Makira (15.4889°S, 049.0785°E, 607 m a.s.l.) GoogleMaps ; ZSM 484/2016 ( ZCMV 15180 ), adult male, collected by M.D. Scherz, A. Rakotoarison, M. Bletz, M. Vences, and J. Razafindraibe on 17 November 2016 at Camp 3 ‘ Simpona’ on the Marojejy Massif (ca 14.4366°S, ca 049.7434°E, ca 1325 m a.s.l.) GoogleMaps .
Diagnosis.— Mantidactylus jonasi sp. nov. is a member of the M. betsileanus clade as revealed by the phylogenomic analysis, and sister to the monophyletic group containing M. betsileanus and M. incognitus . See Table 4 View TABLE 4 for a list of diagnostic morphological characters.
The combination of a relatively small body size in males (SVL 22–25 mm), slightly to moderately tubercular dorsal skin with distinct continuous dorsolateral ridges, reduced webbing (one phalanx of fifth toe free of web), absence of white spots on flanks, presence of a white marking on snout tip, and advertisement call consisting of a single, long pulsed note distinguishes M. jonasi sp. nov. from species of all other clades. Within the M. betsileanus clade, the new species has long been confused with M. betsileanus from which it cannot be reliably distinguished based on morphology, although it may have a tendency towards a somewhat more tubercular dorsal skin than M. betsileanus ( Figs 40 View FIGURE 40 vs 34), and a slightly smaller body size. It can be distinguished from M. betsileanus by fewer pulses per note, and lower pulse repetition rate, in advertisement calls ( Table 4 View TABLE 4 ). Mantidactylus jonasi sp. nov. is distinguished from M. noralottae by smaller size of males and presence of a distinct white marking on snout; from M. tripunctatus by a smaller size, and by fewer pulses per note and a lower pulse repetition rate, in advertisement calls; and from M. incognitus by absence of distinct dorsal ridges and less strongly expressed supraocular tubercles. For a detailed distinction from other new species described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. jonasi sp. nov. in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Description of the holotype.—Adult male in good state of preservation ( Fig. 33 View FIGURE 33 ). Some muscle tissue removed from left thigh. Body relatively slender. Head as wide as body. Snout rounded in dorsal and lateral views. Nostrils directed laterally, slightly protuberant, nearer to tip of snout than to eye. Canthus rostralis weakly recognisable, slightly concave; loreal region slightly concave. Tympanum distinct, large, wider than high, horizontal diameter of tympanum 93% of horizontal eye diameter. Supratympanic fold poorly recognisable, basically identical with outer edge of tympanum and tightly following it. Tongue ovoid, distinctly bifid.
Maxillary teeth present. Vomerine teeth form two rounded aggregations, positioned posterolateral to choanae.
Choanae rounded. Subarticular tubercles single. Inner and outer metacarpal tubercles present. Fingers without webbing. Relative length of fingers: I<II<IV<III. Finger discs slightly enlarged. Nuptial pads absent. Foot longer than tibia (112%). Lateral metatarsalia separated. Inner metatarsal tubercle present. Outer metatarsal tubercle not clearly recognisable. Webbing formula: 1(traces), 2i(traces), 2e(1.5), 3i(2), 3e(2), 4i(2.25), 4e(2.5), 5(1). Relative length of toes: I<II<V<III<IV. Skin on the upper surface smooth with weakly recognisable dorsolateral folds and some granules on flanks (in life, dorsolateral folds distinct and more granular on dorsum). Ventral side smooth. Femoral glands small and at substantial distance from cloaca, with a distinct distal ulcerous macrogland; no clearly recognisable proximal granular gland field.
Colour in preservative: dorsally dark brown with some variation in tone on central dorsum. Ventrally of dorsolateral folds, with slightly darker brown which then sharply borders on a light beige flank colour with some remains of reddish colour. Frenal area light with some dark markings. Anterior dorsal surface of head lighter than dorsum. A distinct and contrasting white patch on tip of snout. Limbs with dark crossbands. Ventrally light beige, throat and chest dirty brown with some light-dark pattern on chest and a white median line on throat. Upper lip ventrally dark brown with white spots. Colour in life similar to preservative but more contrasting colours; the light colouration on flanks and sides of head was orange.
Variation.—Variation in measurements is given in Table 7. See Fig. 40 View FIGURE 40 for colouration in life and its variation. No females were available to us to assess sexual dimorphism. Femoral glands in life often more distinct than in M. betsileanus , with clearly recognisable proximal granular gland field, not conspicuously coloured.
Natural history.—Regularly observed along different kinds of running waters, in primary and degraded rainforest. Males were heard calling from the edges of rainforest streams at night. Despite a superficial similarity with M. betsileanus , this species appears to occur less often in swamps and water bodies such as rice fields.
The holotype was collected beside a spring or seep in disturbed forest with little understory, only a few metres from a larger river. Nearby, a nest containing a cluster of eggs and a frog (presumed to be a guarding parent) was observed ~ 1 m above the water line, along the bank of the spring (MDS, pers. obs.).
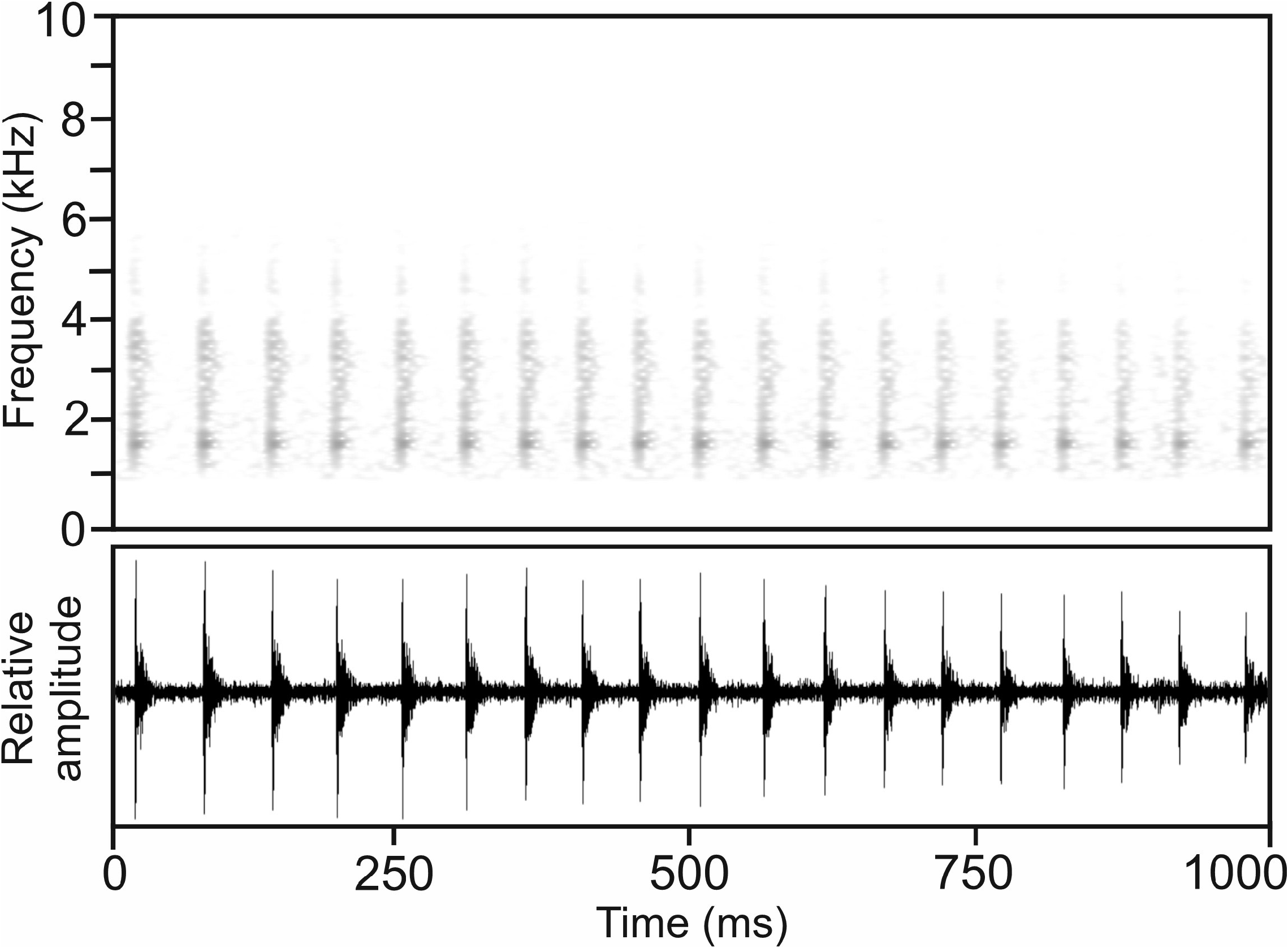
Calls.—Calls recorded on 9 January 2016, 23:37 h, at Ampotsidy in the Bealanana District from the holotype, air temperature not measured but likely below 20°C, were moderately motivated and consisted of a pulsed note, with pulse repetition rate increasing within the note from its beginning to its end. Numerical call parameters of six analysed calls were as follows: call duration (= note duration) 1589–2120 ms (1899.6 ± 192.9 ms); 44–50 pulses per note (46.3 ± 2.6); pulse duration 4–6 ms (5.1 ± 0.5 ms); pulse repetition rate within notes 13.8–30.2 pulses/s (23.1 ± 6.8); dominant frequency 1464–1550 Hz (1498 ± 33 Hz); prevalent bandwidth 800–4800 Hz.
Calls recorded on 20 June 2009, 20:00 h,at the western slope of the Makira Massif, air temperature estimated at 20–25°C, seemed to be reasonably motivated and generally agreed with calls from Ampotsidy, Bealanana district, described above. The call consisted of a regularly pulsed note ( Fig. 41 View FIGURE 41 ) of rather variable duration. Numerical call parameters of 14 analysed calls were as follows: call duration (= note duration) 1060–2773 ms (1919.9 ± 446.1 ms); 19–45 pulses per note (32.9 ± 7.6); pulse duration 6–12 ms (9.1 ± 1.6 ms); pulse repetition rate within notes 16.6–18.9 pulses/s (17.9 ± 1.0); dominant frequency 1644–1755 Hz (1681 ± 45 Hz); prevalent bandwidth 900–5500 Hz; call repetition rate (= note repetition rate) ca 3–5 calls/min.
Calls recorded on 7 June 2010 at Ambatoria, Tsaratanàna, air temperature estimated 20–25°C, also showed a rather low pulse repetition rate, comparable to calls from Makira West and Ampotsidy. Numerical call parameters of six analysed calls were as follows: call duration (= note duration) 2152–2895 ms (2466.3 ± 255.6 ms); 53–72 pulses per note (61.7 ± 7.5); pulse duration 4–6 ms (4.2 ± 0.5 ms); pulse repetition rate within notes 14.7–29.9 pulses/s (25.3 ± 5.4); dominant frequency 1217–1415 Hz (1280 ± 68 Hz), with a second frequency peak of almost identical call energy at ca 2700–2800 Hz; prevalent bandwidth 800–4800 Hz; call repetition rate (= note repetition rate) ca 4–5 calls/min.
Tadpoles.— The tadpole of this species has not been described.
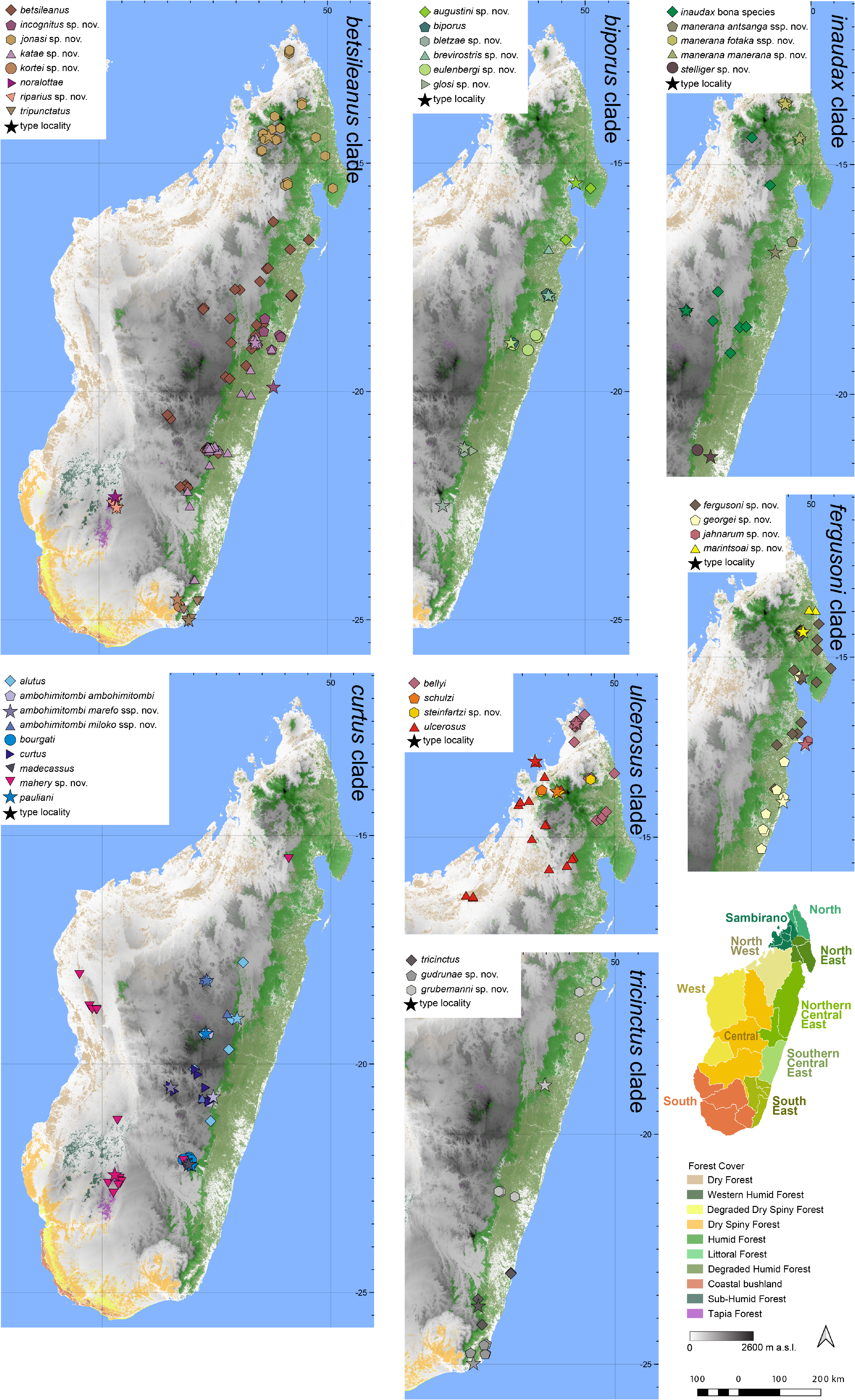
Distribution.— Widespread in northern Madagascar ( Fig. 7 View FIGURE 7 ). This species is known from Ampotsidy, Ambodivohitra,Ampofoko,Andranonafindra,Antambato, Bemanevika, Makira, Mangindrano, Marojejy, Masoala, Montagne d’Ambre, Sorata, and Tsaratanàna. Elevation range: 411–1538 m a.s.l.
Etymology.—L. Rancilhac wishes to dedicate this species to his brother, Jonas Rancilhac, in recognition of his personal support.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |