
Mantidactylus georgei, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504395 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FF5F-FF48-4CB1-4DAF49987ACE |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus georgei |
| status |
sp. nov. |
Mantidactylus georgei sp. nov.
Identity and justification.—This lineage of the M.fergusoni cladehaspreviouslybeenconsideredasconfirmedcandidate species M. sp. 36 by Vieites et al. (2009) and M. sp. Ca36 by Perl et al. (2014). It was depicted as ‘ Mantidactylus sp. aff. betsileanus “ Toamasina ”’ by Glaw and Vences (2007). It was additionally referred to as Mantidactylus sp. aff. betsileanus [Ca AY848260 View Materials ] by Rosa et al. (2011, 2012). It consists of frogs that are bioacoustically somewhat similar to M. betsileanus , but strongly differ in genetics and in various aspects of morphology and advertisement calls ( Table 4 View TABLE 4 ). Mantidactylus georgei sp. nov. forms a clade with another lineage from Maroantsetra considered as confirmed candidate species M. sp. 35 by Vieites et al. (2009) and M. sp. Ca35 by Perl et al. (2014), which we here consider to be a deep conspecific lineage. Due to the high genetic divergence and distinct bioacoustic differentiation of M. georgei sp. nov. to other species in the M. fergusoni clade, its status as distinct species is out of question.
Holotype.— ZSM 455/2005 ( ZCMV 806 ), adult male, collected on 23 February 2004 by M. Vences ca 10 km north of Toamasina (coordinates not taken; coordinates of Toamasina city: 18.167°S, 049.383°E, 22 m a.s.l.), Antsinanana Region , Madagascar.A 16S barcode sequence of the holotype is available from GenBank (accession AY848261 View Materials ). GoogleMaps
Paratypes.—A total of six paratypes: ZSM 456/2005 ( ZCMV 807 ), an adult male with similar collection data to the holotype; MRSN A6213 , adult male, and MRSN A6223 , adult female, both from Betampona, Maintimbato (17.8940°S, 49.2283°E), collected on 21 February 2007 by G.M. Rosa and J. Noël; GoogleMaps MRSN A6341 , adult male, with same locality and collectors as previous specimens but collected on 17 November 2007; GoogleMaps MRSN A6217 and MRSN A6599 , adult males, both collected at Betampona, Rendrirendry (17.9186°S, 49.2103°E), by G.M. Rosa and J. Noël, on 23 March 2007 and 30 November 2007, respectively GoogleMaps .
Additional material.— ZSM 454/2005 ( ZCMV 803 ) an adult male from a genetically divergent population, collected by M. Vences on 24 February 2004 at Maroantsetra (15.4456°S, 049.7375°E, 10 m a.s.l.). GoogleMaps ZMA 19496 ( FGMV 2002.2259 , adult female, collected near Tomasina by M. Vences on 10 February 2003 (specimen not DNA barcoded).
Diagnosis.— Mantidacytlus georgei sp. nov. is a member of the M. fergusoni clade as revealed by the phylogenomic analysis, and splits from a basal node of this clade. See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of small to moderate body size in males (SVL up to 28–31 mm), moderately tubercular dorsal skin, large tympanum size in males (13– 15% of SVL), absence of white spots on flanks, presence of a white marking on the snout tip of many individuals (especially males), and advertisement call consisting of a single, relatively long and pulsed note distinguishes M. georgei sp. nov. from most species of the other clades ( Table 4 View TABLE 4 ). Two species from the M. ulcerosus clade ( M. ulcerosus and M. bellyi ) can be morphologically similar, but they occur in the Sambirano and North West regions, and exhibit higher pulse repetition rates in advertisement calls ( Table 4 View TABLE 4 ). M. georgei sp. nov. shows similarities to species of the M. betsileanus clade but in general it has a more tubercular dorsum and differs from most of these by advertisement call structure ( Table 4 View TABLE 4 ); furthermore, sympatry with species of the M. betsileanus clade is rare, and is only known from Betampona where the new species co-occurs with M. betsileanus . Within the M. fergusoni clade, it strongly differs from M. fergusoni in advertisement call structure, and by the more common presence of a white marking on the snout tip. For a distinction from other new species in the M. fergusoni clade, see below. A full list of molecular diagnostic sites in the 16S gene of M. georgei sp. nov. in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Description of the holotype. —Adult male in good state of preservation ( Fig. 48 View FIGURE 48 ). Tissue sample taken ventrally from right thigh. Remaining (left) femoral gland partly detached to examine its structure internally. Second and third finger of left hand mutilated. Body rather slender. Head as wide as body. Snout rounded. Nostrils directed laterally, slightly protuberant. Nostrils nearer to tip of the snout than to eye. Canthus rostralis weak, slightly concave. Loreal region weakly concave.
Tympanum distinct, large, elliptical, wider than high, its diameter 96% of eye diameter. Supratympanic fold distinct, beginning straight, with a rather distinct, angular bend midway towards insertion of forelimb. Tongue ovoid, distinctly posteriorly bifid. Maxillary teeth present.
Vomerine teeth present in two rounded aggregations, positioned posterolateral to choanae. Choanae rounded.
Subarticular tubercles single. Outer metacarpal tubercle present, inner metacarpal tubercle present. Fingers without webbing. Relative length of fingers: I=II<IV<III.
Finger discs slightly enlarged. Nuptial pads absent. Foot longer than tibia (103%). Lateral metatarsalia separated.
Inner metatarsal tubercle present. Outer metatarsal tubercle small but recognisable. Webbing formula: 1(1), 2i(1), 2e(1), 3i(2), 3e(1), 4i(2), 4e(2), 5(0.5). Relative length of toes: I<II<V<III<IV. Skin on the upper surface with granules, many of which form irregular longitudinal ridges. Ventral side smooth. Femoral glands present, with a distinct distal ulcerous macrogland and a moderately expressed proximal granular gland field.
Colour in preservative: dorsum brown, with indistinct irregular darker markings. A dark brown band between eyes is attached to a grey band on the head surface.
Forelimbs brown with poorly defined darker markings.
Hindlimbs brown with darker crossbands. Inguinal region without scattered whitish spots. Snout tip with a distinct light dot. Venter beige, throat and belly with very little brown mottling on thorax. Lower lip ventrally with alternating light and brown spots on the one side, on the other side uniformly beige-brown. Toe discs dark.
Variation.—Variation in measurements is given in Table 8. See Fig. 51 View FIGURE 51 for colouration in life and its variation.
No females available to assess sexual dimorphism.Femoral glands in life are rather distinct and prominent in males, consisting mostly of a large distal ulcerous macrogland and with only weakly recognisable proximal granular gland field, often with a somewhat yellowish colour; small, rudimentary glands recognisable in females.
Natural history.—Both near Toamasina and Maroantsetra, calling males were found at night from the shores of small streams and swampy areas next to ricefields, in heavily degraded secondary forest habitat. In Betampona and Ivoloina this species is commontly found along stream banks. In Ivoloina M. georgei is the only Brygoomantis species that can be found in the area, while in Betampona it occurs (often syntopically) with four Brygoomantis species. In Betampona it has been recorded in all campsites except Sahabefoza and Sahembendrana (which correspond to the campsites presenting the most pristine conditions).
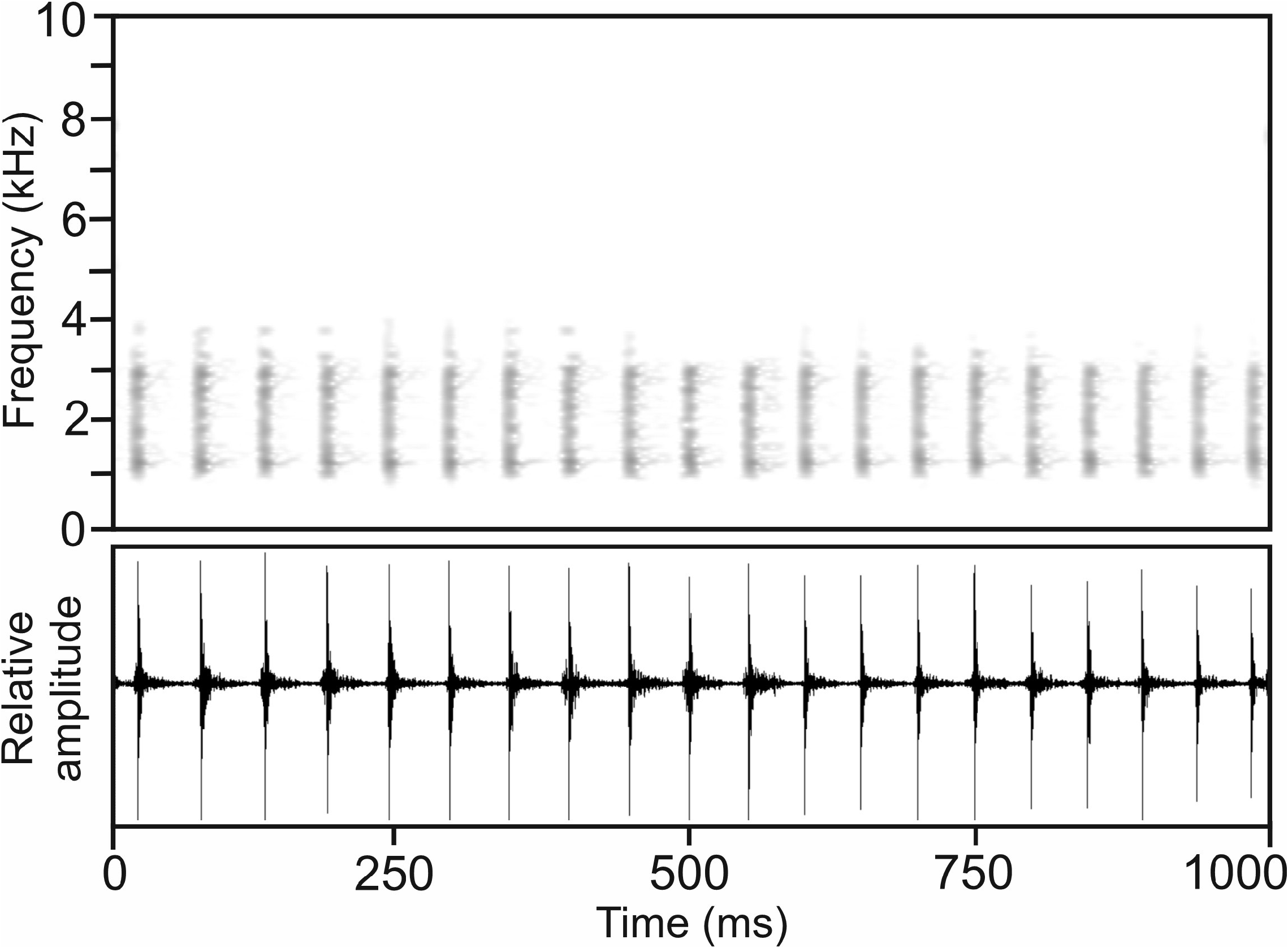
Calls.— The advertisement call of the holotype of M. georgei recorded on 29 February 2004 a few km north of Toamasina, 25.2°C air temperature ( Vences et al. 2006: CD2, track 66), consisted of a long to very long, regularly pulsed note ( Fig. 52 View FIGURE 52 ). Notes exhibited slight amplitude modulation, with call energy decreasing towards the note’s end. Numerical parameters of 11 analysed calls were as follows: call duration (= note duration) 1425–3206 ms (2532.7 ± 527.4 ms); 26–64 pulses per note (46.9 ± 11.3); pulse duration 5–12 ms (7.9 ± 2.1 ms); pulse repetition rate within notes 12.7–24.4 pulses/s (18.1 ± 3.3); dominant frequency 1227–1722 Hz (1483 ± 184 Hz), with a second peak of almost identical energy at around 2970–3040 Hz; prevalent bandwidth 900–3400 Hz; call repetition rate not possible to identify with the available recordings.
Calls recorded on 24 February 2004, 21:00 h, at Maroantsetra, 24.1°C air temperature ( Vences et al. 2006: CD2, track 65), very likely correspond to M. georgei and generally agreed with the calls described above, apart from somewhat higher pulse repetition rate and slightly higher number of pulses per note. Numerical parameters of six analysed calls were as follows: call duration (= note duration) 2470–3149 ms (2826.8 ± 278.7 ms); 51– 85 pulses per note (71.7 ± 14.9); pulse duration 5–10 ms (7.9 ± 0.9 ms); pulse repetition rate within note 17.8–32.6 pulses/s (25.0 ± 4.9); dominant frequency 1260–1345 Hz (1287 ± 34 Hz); prevalent bandwidth 900–3800 Hz; call repetition rate not possible to identify with the available recording.
Additional calls have been recorded at Betampona and present an overall similar structure to the previous descriptions for the species ( Rosa et al. 2011: track 35).
Tadpoles.— The tadpole of this species has not been described.
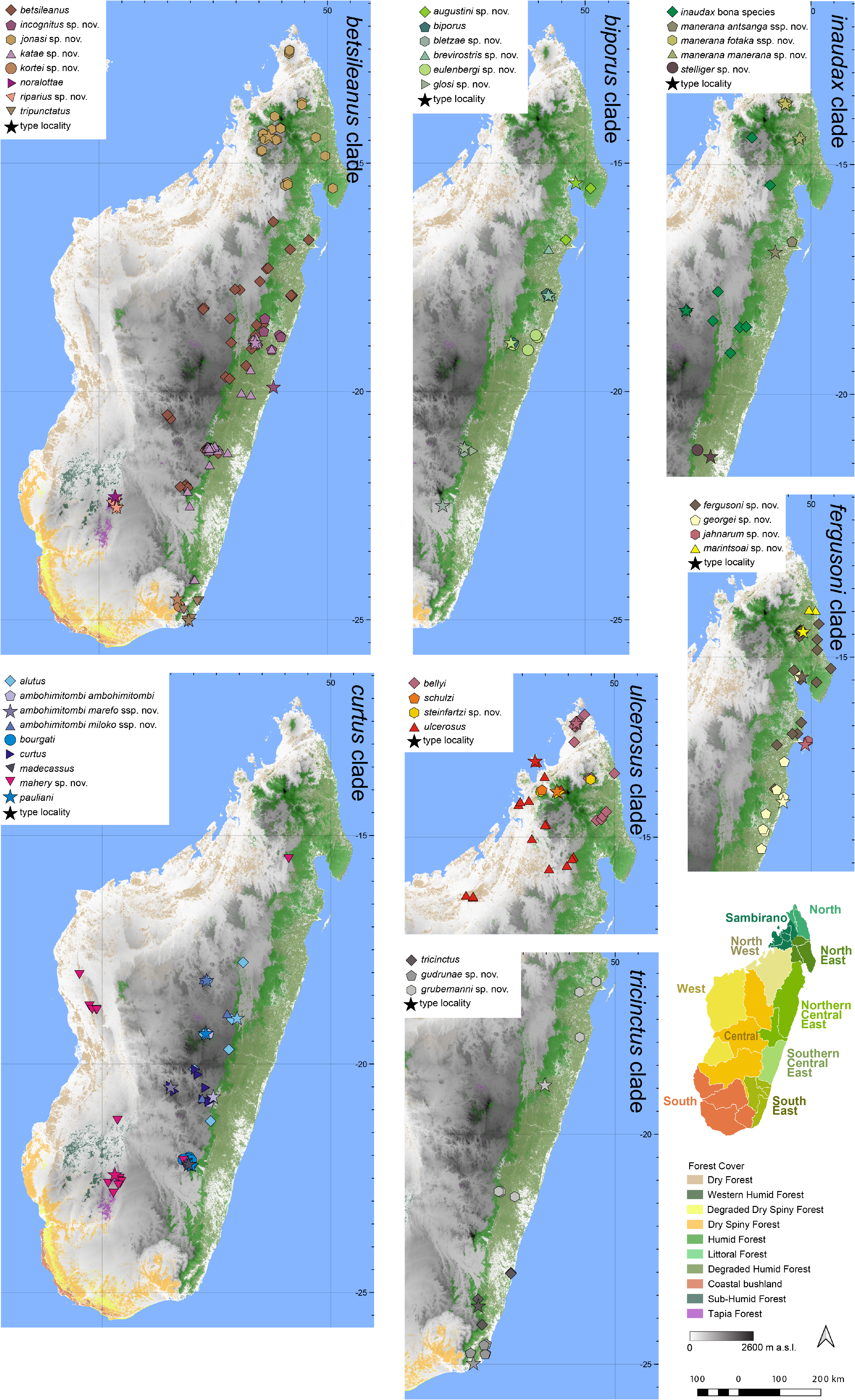
Distribution.— Endemic to low-elevation areas of the Northern Central East of Madagascar ( Fig. 7 View FIGURE 7 ). This species is known from Anivorano Est, Antokotelo, Betampona, Ivoloina, Maroantsetra, Sahafina, Tampolo forest (Analanjirofo), Toamasina, and the vicinity of Vatomandry. Elevation range: 7–517 m a.s.l.
Etymology.—We dedicate this species, which occurs, among other sites along Madagascar’s east coast, in Betampona Strict Nature Reserve, to Georges, our guide on numerous field expeditions in Betampona, in recognition of his dedication to the study of the herpetofauna of Betampona. The species epithet georgei is derived from the English translation of our guide’s name (George) as it thereby becomes easier to pronounce.


| MRSN |
Italy, Torino, Museo Regionale di Scienze Naturali |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |