
Mantidactylus bourgati Guibé, 1974
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504348 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FF95-FF82-4F13-48CF4C2F78EE |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus bourgati Guibé, 1974 |
| status |
|
Mantidactylus bourgati Guibé, 1974 View in CoL
Type material.— Mantidactylus bourgati Guibé, 1974 is based on the holotype MNHN 1972.437 (given in error as 1972.427 in the original publication but correct in the handwritten catalogue) by original designation from ‘ Ambalamarovandana’ , a site in the Andringitra Mountains , and seven paratypes ( MNHN 1972.440 , 1972.449 , 1972.464 , 1972.472 , 1972.476 , 1972.479 , and 1972.491 )
Identity.— Mantidactylus bourgati has long been considered a synonym of M. curtus but was resurrected as a separate species by Glaw and Vences (2006) based primarily on strong genetic divergences. We here provide a 16S sequence obtained by barcode fishing from the holotype MNHN 1972.437 confirming the attribution of the lineage containing all samples of curtus -like stream frogs from the Andringitra Massif to this nomen.
Diagnosis.—A member of the M. curtus clade and sister to M. curtus . See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of relatively large body size of at least up to 41 mm ( Table 5 View TABLE 5 ; larger specimens observed in the field), slightly granular skin, strongly expressed foot webbing with almost fully webbed fifth toe distinguishes this species from species of the other clades. Within the M. curtus clade, M. alutus , M. madecassus and M. pauliani have smaller body sizes ( Table 4 View TABLE 4 ). M. curtus has a very smooth dorsal skin while in M. bourgati the skin is usually somewhat granular. Morphological distinction from M. a. ambohimitombi and M. a. miloko is most difficult, but as far as known, the distributions of these taxa do not overlap with that of M. bourgati . M. bourgati differs further from M. alutus by longer advertisement calls emitted at irregular intervals (vs regular series in M. alutus ). For detailed distinction from new species described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. bourgati in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Variation. —Variation in measurements is given in Table 5 View TABLE 5 . See Fig. 17 View FIGURE 17 for colouration in life and its variation. From our sample, it is unclear whether sexual size dimorphism exists in this species; in the specimens measured, male SVl is 33.0– 38.9 mm (n=2) and female SVL is 32.3–40.0 mm (n=13). Femoral glands in adult males in life are often clearly visible and prominent ( Fig. 17 View FIGURE 17 ) but in preserved specimens, a clear distinction between males and females (which probably also have visible gland rudiments) is often not possible without examination of gonads.
Natural history.—Specimens of M. bourgati are common in and around small slow-flowing montane streams at high elevations in the Andringitra Massif, e.g. on the Andohariana Plateau at 2000 m a.s.l. where they can be found at the edge of the water. They do not seem to occur higher up the massif, e.g. at Cuvette Boby we found only M. madecassus . However, at somewhat lower elevations (e.g. Imaitso forest at ca 1500 m a.s.l.), M. bourgati is also common along large streams running through rainforest. Our call recordings were made from a swamp area next to such a stream where specimens were observed during the day in the shallow water.
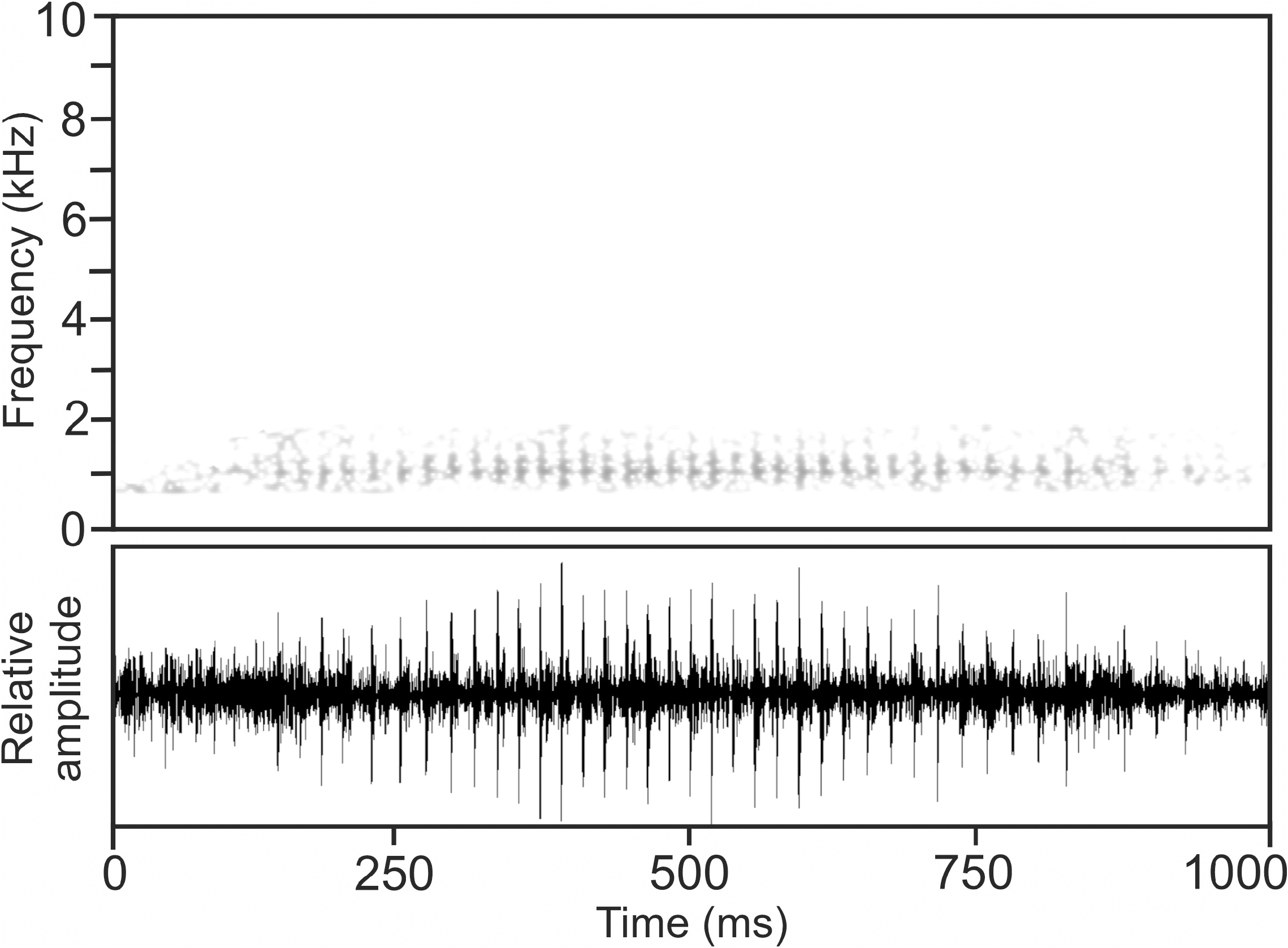
Calls.— The advertisement call of M. bourgati , has been recorded on 16 January 1994, 10:00 h, at Andringitra National Park, in a forest swamp area close to Ambalamarina, 19°C air temperature, but the calling specimens could not be seen, and the attribution to M. bourgati is tentative ( Vences et al. 2006: CD 2, track 77).
The call consists of a regularly pulsed note( Fig.18 View FIGURE 18 ),emitted in series at somewhat irregular intervals. Notes exhibit some amplitude modulation, with amplitude continuously increasing from the beginning, reaching maximum call energy at the middle of the note, before continuously decreasing towards the note’s end. Numerical parameters of four analysed calls are as follows: call duration (= note duration) 756–1020 ms (872.8 ± 112.6 ms); 36–50 pulses per note (43.8 ± 5.9); pulse duration 8–10 ms (8.7 ± 0.8 ms); pulse repetition rate within notes 45.8–54.1 pulses/s (50.8 ± 4.1); dominant frequency 1063–1106 Hz (1087 ± 18 Hz); prevalent bandwidth 880–1740 Hz; call repetition rate (= note repetition rate) within series ca 19–29 calls/min.
Tadpoles.— The tadpole of this species has not been described.
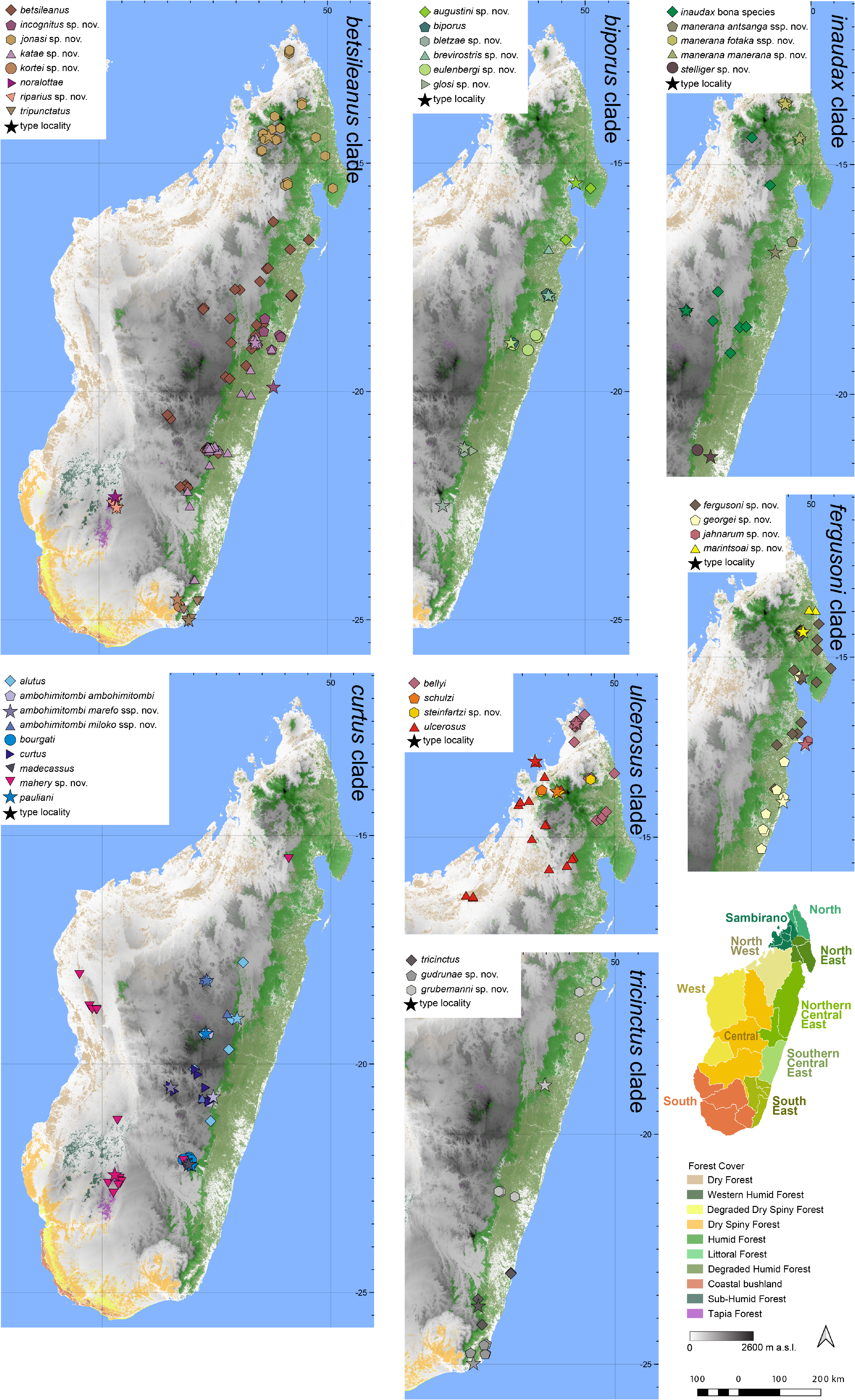
Distribution.— Apparently microendemic to Andringitra and nearby areas ( Fig. 7 View FIGURE 7 ). This species is known from Andringitra (various sites, type locality), Belambo, Fivahona, and Tsaranoro. Elevation range: 940–2488 m a.s.l.
Etymology.—Eponym for ‘Professor [Robert M.] Bourgat of the University of Lomé ( Togo), who collected several frogs during his voyages in Madagascar’(translated from Guibé 1973a).

| MNHN |
France, Paris, Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |