
Mantidactylus augustini, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504415 |
|
persistent identifier |
https://treatment.plazi.org/id/34970990-9D11-4871-AED2-E5CF7466FA2C |
|
taxon LSID |
lsid:zoobank.org:act:34970990-9D11-4871-AED2-E5CF7466FA2C |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus augustini |
| status |
sp. nov. |
Mantidactylus augustini sp. nov.
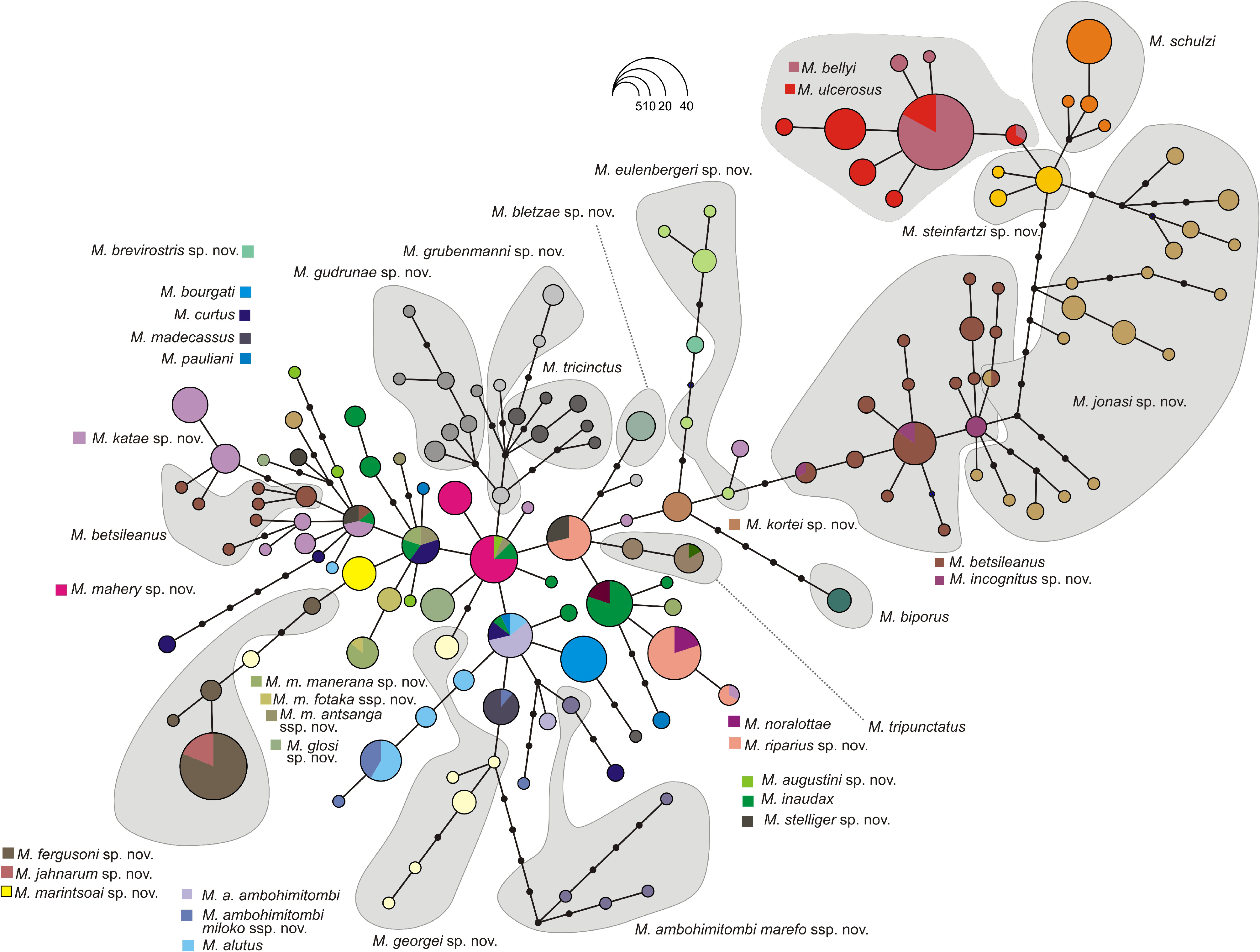
Identity and justification.—This lineage is a member of the M. biporus clade from the lowlands of the North East of Madagascar and has previously been considered as confirmed candidate species M. sp. 22 by Vieites et al. (2009) and M. sp.Ca22by Perl et al. (2014).It was depicted as ‘ Mantidactylus sp. aff. biporus “Andranofotsy”’ by Glaw and Vences (2007). This lineage is sister to the true M. biporus according to our phylogenomic analysis, but is characterized by a high uncorrected pairwise-distance in the 16S rRNA marker (4.9–5.8%). It also is concordantly differentiated in the nuclear Rag-1 gene, not sharing its haplotype with M. biporus ( Fig. 4 View FIGURE 4 ). Moreover, the two lineages differ distinctly in male advertisement call. Based on the concordance of high mitochondrial divergence with nuclear and bioacoustic differentiation, we are convinced it represents a distinct species. A deep conspecific lineage of M. augustini sp. nov. co-occurs with the main lineage in Masoala. As we have only limited data on this lineage, we tentatively include it in our circumscription of this species, but note that it may later transpire to represent another distinct species.
Holotype.— ZSM 122/2002 ( MV 2001.1388 ), adult male, collected by M. Vences on 17 December 2001 at Andranofotsy (wood nearby, 15.4353°S, 049.8439°E, 85 m a.s.l.), Analanjirofo Region , Madagascar. 16S and cox1 barcode sequences of the holotype are available from GenBank (accessions AY848225 View Materials and JN133225 View Materials ). GoogleMaps
Paratypes.—A total of six paratypes: ZSM 740/2009 ( ZCMV 11170 ), possibly female, collected by J.E. Randrianirina on 15 May 2009 at Melivinany ‘S 0I’, Manompana, Forêt de Befanjana (precise coordinates unavailable); GoogleMaps MRSN A3600 ( FN 7678 = ACZC 4904 ), adult female, collected by F. Andreone, and J.E. Randrianirina on 1 December 1998 in Beanjada ‘Corridor 1’, Ambatoledama Corridor, Masoala (ca 15.267°S, ca 049.983°E, ca 1000 m a.s.l.); GoogleMaps MRSN A2905 ( FN 7238 = ACZC 4897 ), possibly female, collected by J.E. Randrainirina on 6November 1998 at Andranobe, Masoala National Park (coordinates unavailable); GoogleMaps MRSN A3540 ( ACZC 4898 ), possibly female, collected by R. Nincheri on 24 July 1993 in Masomihenija forest , Ambodilalono , Masoala peninsula (coordinates unavailable); GoogleMaps MRSN A6740 ( FAZC 14292 = ACZC 4906 ), presumed subadult female, collected by J.E. Randrianirina on 27 April 2008 at Farankaraina (coordinates unavailable); GoogleMaps MRSN A3737 ( FAZC 10009 = ACZC 4905 ), juvenile, collected by F. Andreone and J.E. Randrianirina on 1 December 1999 at ‘Camp 4’, Antsarahan’Ambarato in the Ilampy Corridor , Masoala peninsula (15.3920°S, 050.0470°E, ca 550 m a.s.l.) GoogleMaps .
Additional material. — ZFMK 70481 from Masoala probably belongs to this species but is not included in the paratype series due to the lack of genetic data.
Diagnosis.— M. augustini sp. nov. is a member of the M. biporus clade, sister to M. biporus according to our phylogenomic analysis. See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of a small body size (male SVL 24 mm, female SVL 21–25 mm), rather smooth dorsal skin with weakly expressed dorsolateral ridges in some individuals, large tympanum size in males (13% of SVL), presence of white spots on flanks, absence of a white marking on the snout tip, and a short, pulsed advertisement call emitted in regular series, distinguishes M. augustini sp. nov. from species of the M. betsileanus , M. curtus , M. fergusoni , M. tricinctus , and M. ulcerosus clades. Mantidactylus inaudax ( M. inaudax clade) is morphologically similar but appears to have shorter hindlimbs, less pulses per note, and higher pulse repetition rate in advertisement calls. M. augustini sp. nov. is distinguished from its sister species, M. biporus , by smaller body size, larger tympanum, longer hindlimbs, more pulses per note and a lower pulse rate in advertisement calls ( Table 4 View TABLE 4 ), as well as a higher dominant frequency (1263–1356 Hz vs 832–997 Hz), and a lower call repetition rate (200–230 vs 300–360 calls/ min). For a distinction from the other (all new) species in the M. biporus , M. stelliger and M. inaudax clades, see the diagnoses in the respective species accounts below.
A full list of molecular diagnostic sites in the 16S gene of M. augustini sp. nov. in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Description of the holotype. Adult male in moderate state of preservation ( Fig. 62 View FIGURE 62 ). Left foot missing (taken as tissue sample), femoral glands partly detached for examination in internal view. Body rather stout. Head wider than body. Snout rounded. Nostrils directed laterally, slightly protuberant. Nostrils nearer to tip of the snout than to eye. Canthus rostralis weak, slightly concave. Loreal region weakly concave.Tympanum distinct, large, rounded, diameter 82% of eye diameter. Supratympanic fold closely following outer edge of tympanum, not clearly recognisable as separate structure in preserved specimen. Tongue ovoid, distinctly posteriorly bifid. Maxillary teeth present. Vomerine teeth present in two rounded aggregations, positioned posterolateral to choanae. Choanae rounded. Subarticular tubercles single. Outer metacarpal tubercle recognisable, inner metacarpal tubercle present. Fingers without webbing. Relative length of fingers: I<II<IV<III. Finger discs slightly enlarged. Nuptial pads absent. Foot longer than tibia (111%). Lateral metatarsalia separated. Inner metatarsal tubercle present. Outer metatarsal tubercle not recognisable. Webbing formula: 1(1), 2i(1.5), 2e(1), 3i(2), 3e(1.5), 4i(3), 4e(3), 5(1). Relative length of toes: I<II<V<III<IV. Skin on the upper surface smooth. Ventral side smooth. Femoral glands present, relatively small, consisting of a distal ulcerous macrogland; no proximal granular gland field recognisable.
Colour in preservative: dorsum brown, with indistinct irregular darker and lighter markings and a light vertebral band. Forelimbs brown with darker markings. Hindlimbs brown with indistinct darker crossbands. Inguinal region with few whitish spots. Snout tip without a light dot. Venter uniformly beige, throat darker than belly with brown mottling. Lower lip with distinct alternating light and brown spots. Toe discs dark. Toes light and dark striped.
Colour in life: dorsum dark brown with indistinct darker markings. Similar to colour in preservative. Forelimbs brown with indistinct darker markings; hindlimbs with dark crossbands. Venter uniformly beige, throat with distinct brown mottling. A beige vertebral band is present. On the flanks few white spots present. Skin on the back smooth.
Variation.—Variation in measurements is given in Table 10. See Fig. 65 View FIGURE 65 for colouration in life and its variation. Data on sexual size dimorphism is inconclusive, and our sample size is small (confirmed male SVL 23.9 mm [n = 1] vs confirmed female SVL 20.9–25.0 mm [n = 3]). There does not seem to be a clear size dimorphism in tympanum diameter (HTD/ED ratio is 68–92% in females, 82% in the male). Femoral glands of the male in life were not very strongly expressed; small gland rudiments are recognisable also in a female ( Fig. 65c View FIGURE 65 ).
Natural history.—At the type locality, calling males were observed at night from the shore of a very shallow running water of a spring in rainforest.
)
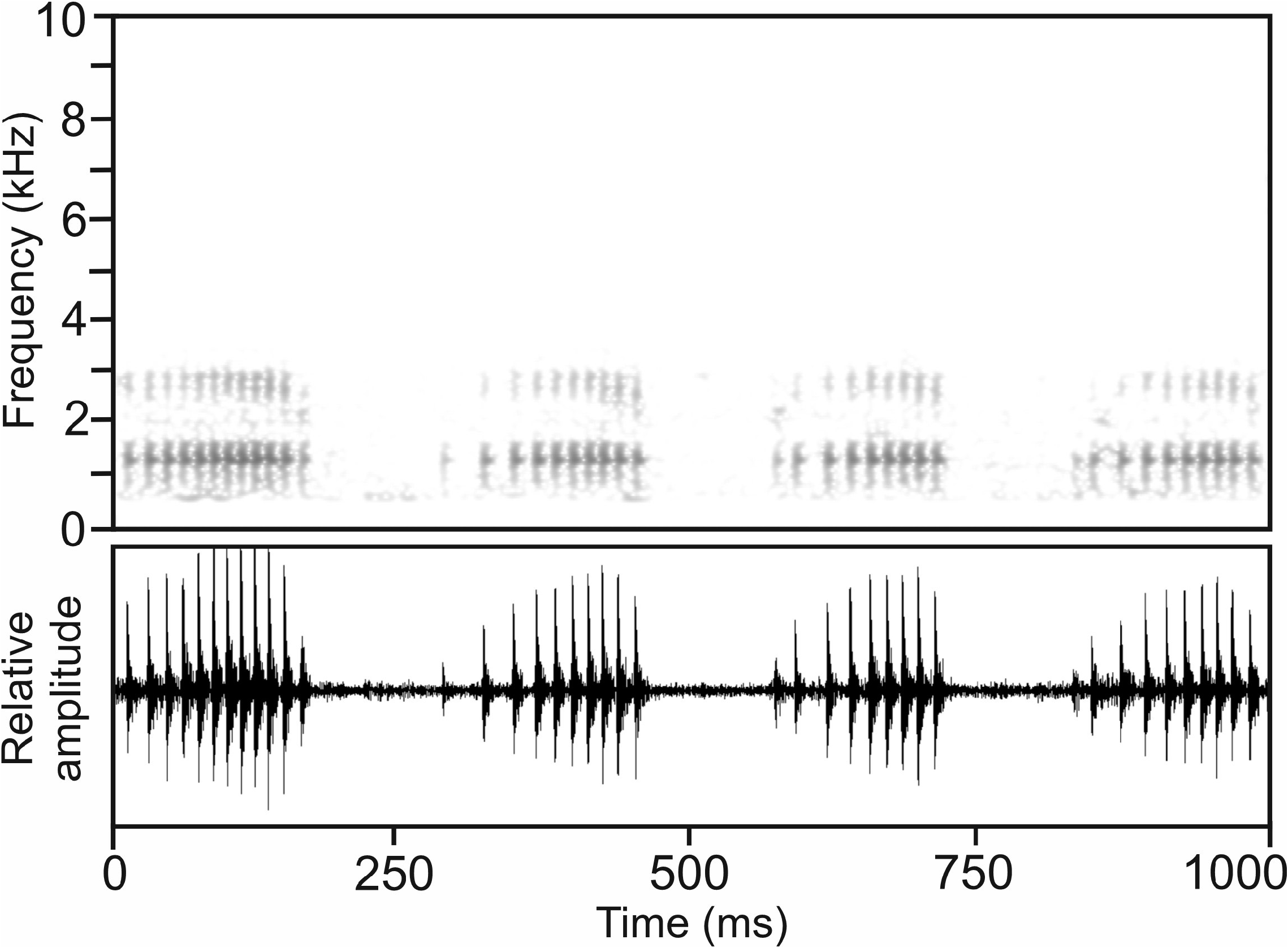
Calls.—The advertisement call of M. augustini , recorded on 16 December 2001 near Andranofotsy, 25.4°C air temperature ( Vences et al. 2006: CD2, track 72), consisted of a short, pulsed note, emitted in regular series at fast succession ( Fig. 66 View FIGURE 66 ). Pulse repetition rate was distinctly lower at the beginning of calls and increased after approximately one half of the call’s duration. Amplitude modulation was present, with relative amplitude increasing from the beginning of the call, reaching its maximum at the last quarter of the call’s duration. Numerical parameters of seven analysed calls were as follows: call duration (= note duration) 154–236 ms (178.8 ± 26.6 ms); 9–15 pulses per note (10.4 ± 2.1); pulse duration 7–12 ms (9.1 ± 1.6); pulse repetition rate within notes 44.4–71.4 pulses/s (59.0 ± 10.3); dominant frequency 1263–1356 Hz (1304 ± 39 Hz); prevalent bandwidth 700–3150 Hz; call repetition rate (= note repetition rate) within regular series ca 200– 230 calls/min.
Tadpoles.— The tadpole of this species has not been described.
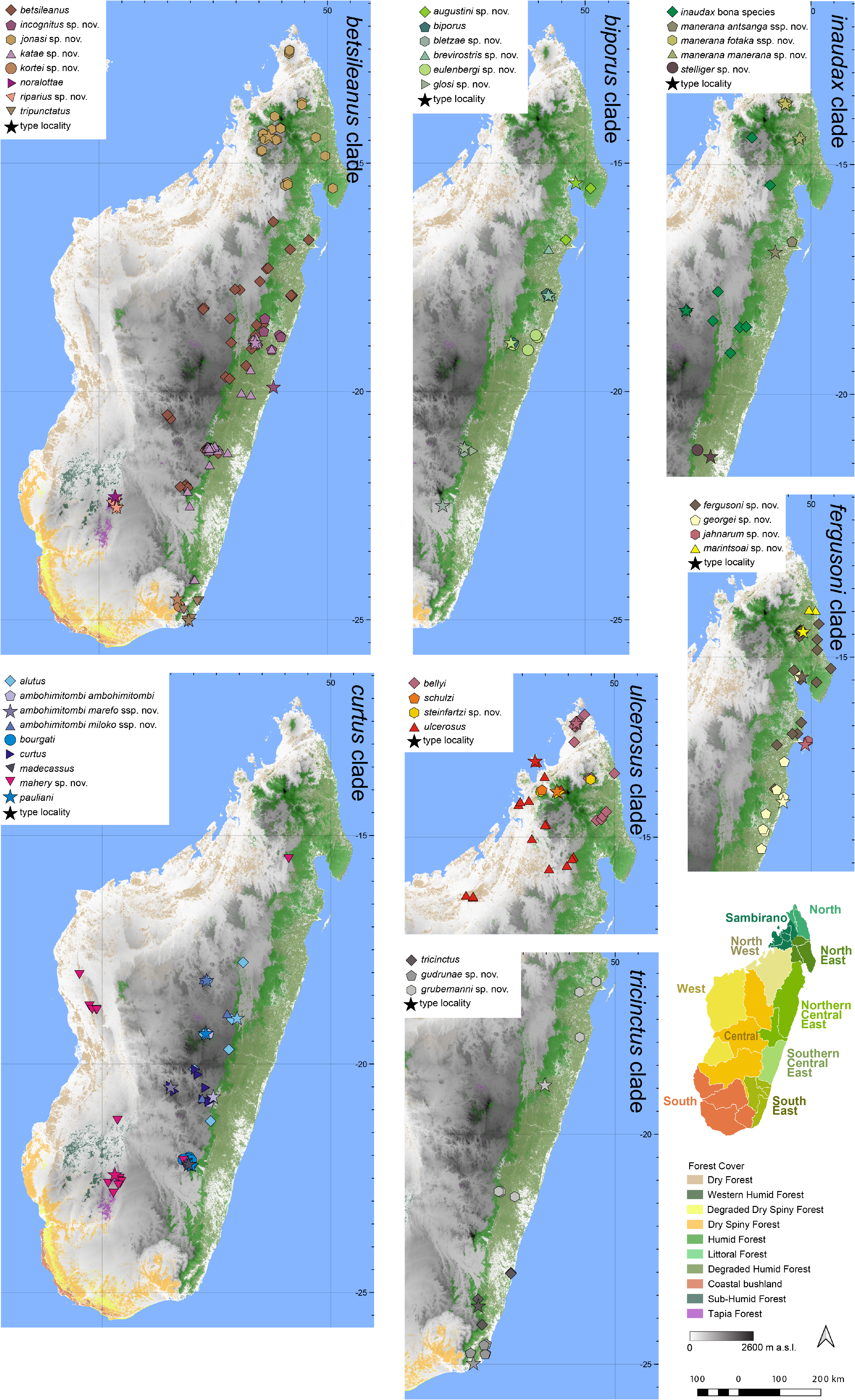
Distribution.— Endemic to the North East of Madagascar ( Fig. 7 View FIGURE 7 ). This species is known from Masoala, Manompana (Befanjana), Antsahataloka, and Andranofotsy. Elevation range: Ranging from near sea level (85 m a.s.l.) to ~ 1000 m a.s.l.
Etymology.—We dedicate this species to Augustin Sarovy, an excellent musician, guide, and ecologist from Maroantsetra, whose help was crucial to collect the holotype of this new species and to record its call.


| MRSN |
Italy, Torino, Museo Regionale di Scienze Naturali |
| ZFMK |
Germany, Bonn, Zoologische Forschungsinstitut und Museum "Alexander Koenig" |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |