
Mantidactylus ambohimitombi miloko, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504346 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FF94-FF87-4CB1-4C0F49827CAE |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus ambohimitombi miloko |
| status |
subsp. nov. |
Mantidactylus ambohimitombi miloko ssp. nov.
Identity and justification.—This lineage of the M. curtus clade was considered as confirmed candidate species M. sp. 18 by Vieites et al. (2009) due to its mitochondrial divergence in concert with slight but distinct differences in colour pattern, and as M. sp. Ca18 by Perl et al. (2014). It was depicted as ‘ Mantidactylus sp. aff. curtus “Ambohitantely”’ by Glaw and Vences (2007). We here consider this lineage provisionally as a subspecies of M. ambohimitombi based on the following rationale: (i) it belongs to the same general mitochondrial lineage as the nominal subspecies in the 16S tree, and the two are also closely related in the phylogenomic tree; (ii) morphologically, the two lineages are similar to each other, except for a somewhat more distinct dorsal pattern in M. a. miloko ssp. nov.; (iii) based on two samples in our mitochondrial tree, RJS 1877 and ACZC 4254, a very similar mitochondrial haplotype to the one from Ambohitantely also occurs at Ankaratra, suggesting the possibility of past or ongoing gene flow between localities or mitochondrial introgression;and (iv)based on sequences of specimens APR 10803, APR 10638, and APR 10663, the lineage also occurs in Angavokely and Ankazomivady, thus rather close to Ankaratra, suggesting the two lineages may be parapatric and could hypothetically have a hybrid zone. Since the status of the Ambohitantely population as a fully isolated evolutionary lineage is thus not fully verifiable with the data at hand, we consider the status as a subspecies of M. ambohimitombi to be adequate.
Holotype.— ZSM 219/2005 (FGZC 2143), adult male, collected by M. Vences, L. du Preez, P. Bora, L. Raharivololoniaina, R.D. Randrianiaina, T. Razafindraibe, E. Randriamitso on 18 January 2005 at Ambohitantely Special Reserve , ‘Jardin Botanique’, at a site about 500 m fromthegeographicalcoordinates 18.1725°S, 047.2768°E, 1580 m a.s.l., Analamanga Region, Madagascar. GoogleMaps A 16S barcode sequence of the holotype was obtained in this study and was included in the analysis.
Paratype.—A single paratype: ZSM 237/2005 (FGZC 2172), adult female, with the same collection data as holotype.
Additional material. —The following specimens probably belong to this taxon but are only assigned tentatively and not designated as paratypes because no molecular data is available: ZMA 6859 (two specimens with field numbers 675 and 676) and ZMA 6860 (six specimens with field numbers 910–914, 917), collected by R.M.A. Blommers-Schl ̂sser in 1972 at 1500 m ( ZMA 6859 ) and 2200 m a.s.l. ( ZMA 6860 ) at Tampoketsa d’Ankazobe .
Diagnosis.— Mantidactylus ambohimitombi miloko is a member of the M. curtus clade, and the sister group of M. a. ambohimitombi + M. a. marefo. Morphologically, it is very similar to M. a. ambohimitombi . See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of relatively large body size of up to 49 mm, slightly granular skin without clearly defined dorsolateral ridges, and small tympanum diameter of a maximum of 10% of SVL in males, distinguishes M. a. miloko ssp. nov. from species of the other clades. Within the M. curtus clade, M. alutus , M. madecassus and M. pauliani have smaller body sizes and are distinguished by either a usually shorter snout ( M. madecassus , M. pauliani , M. a. marefo), or presence of rather distinct dorsolateral ridges ( M. alutus ) ( Table 4 View TABLE 4 ); M. curtus usually has a smoother skin and a somewhat shorter snout; M. bourgati is morphologically very similar but appears to occur only on the Andringitra Massif. Compared to other subspecies of M. ambohimitombi , the new subspecies differs from M. a. marefo by its distinct dorsal pattern, lack of bluish colour around the eye, and more pointed snout; and from M. a. ambohimitombi by the usually more distinct dorsal pattern. For detailed distinction from other new species and subspecies described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. a. miloko in pairwise comparisons to all other Brygoomantis species and subspecies is provided as Supplementary appendix.
Description of the holotype.—Adult male in good state of preservation ( Fig. 9 View FIGURE 9 ). Tissue sample taken ventrally from right thigh. Femoral gland partly detached to examine their structure internally. Body rather slender.
Head slightly wider than body. Snout rounded in dorsal and lateral view. Nostrils directed dorsolaterally, slightly protuberant. Nostrils nearer to tip of the snout than to eye. Canthus rostralis almost not recognisable, slightly concave. Loreal region slightly concave. Tympanum distinct, elliptical, wider than high, its diameter 78% of eye diameter. Supratympanic fold distinct, beginning straight, with a distinct, angular 90° bend at the posterior edge of tympanum towards insertion of forelimb. Tongue ovoid, distinctly posteriorly bifid. Maxillary teeth present. Vomerine teeth distinct in rounded aggregations, positioned posterolateral to choanae. Choanae rounded.
Subarticular tubercles single. Outer metacarpal tubercle present, inner metacarpal tubercle present. Fingers without webbing. Relative length of fingers: I=II<IV<III. Finger discs minimally enlarged. Nuptial pads absent. Foot longer than tibia (113%). Lateral metatarsalia separated.
Inner metatarsal tubercle present. Outer metatarsal tubercle small but recognisable. Webbing formula: 1(0.25), 2i(1), 2e(0.5), 3i(1.5), 3e(1), 4i(2), 4e(2), 5(0.75).
Relative length of toes: I<II<V=III<IV. Skin on the upper surface smooth, with some granules on flanks, and some longitudinal tubercles forming interrupted and weakly expressed dorsolateral folds.Ventral side smooth. Femoral glands distinct but relatively small, with a distal ulcerous macrogland consisting of at least eight large granules and an external central depression, and a weakly expressed proximal granular gland field visible in internal view.
Colour in preservative: light brown dorsally with large and contrasted dark brown patches all over the dorsal surface, distinct dark crossbands on limbs, and a light frenal stripe with some dark markings on the upper lip.
Ventrally uniformly light grey with alternating dark-light pattern ventrally on the lower lip. Colour in life similar to that in preservative, but more contrasted ( Fig. 16 View FIGURE 16 ).
Variation. —Variation in measurements is given in Table 5 View TABLE 5 . Too few specimens have been sexed to assess the degree of sexual size dimorphism. Femoral glands distinct and large (but not differing in colour from surrounding ventral skin of thigh) in the male holotype ( Fig. 16 View FIGURE 16 ). In contrast, specimens from the ZMA collection are difficult to sex externally as femoral glands are often indistinct, as seems to be typical for several taxa in the M. curtus clade, possibly due to seasonal effects.
Natural history.— Specimens were collected around clean highland streams running in open areas between forest fragments. Specimens at Angavokely and Ankazomivady that appear to belong to this taxon based on mitochondrial DNA were found on wet rocks along slow-moving parts of streams.
Calls.—The call of this subspecies has not been recorded.
Tadpoles.— Probably reported from Angavokely by Blommers-Schl̂sser (1979)..
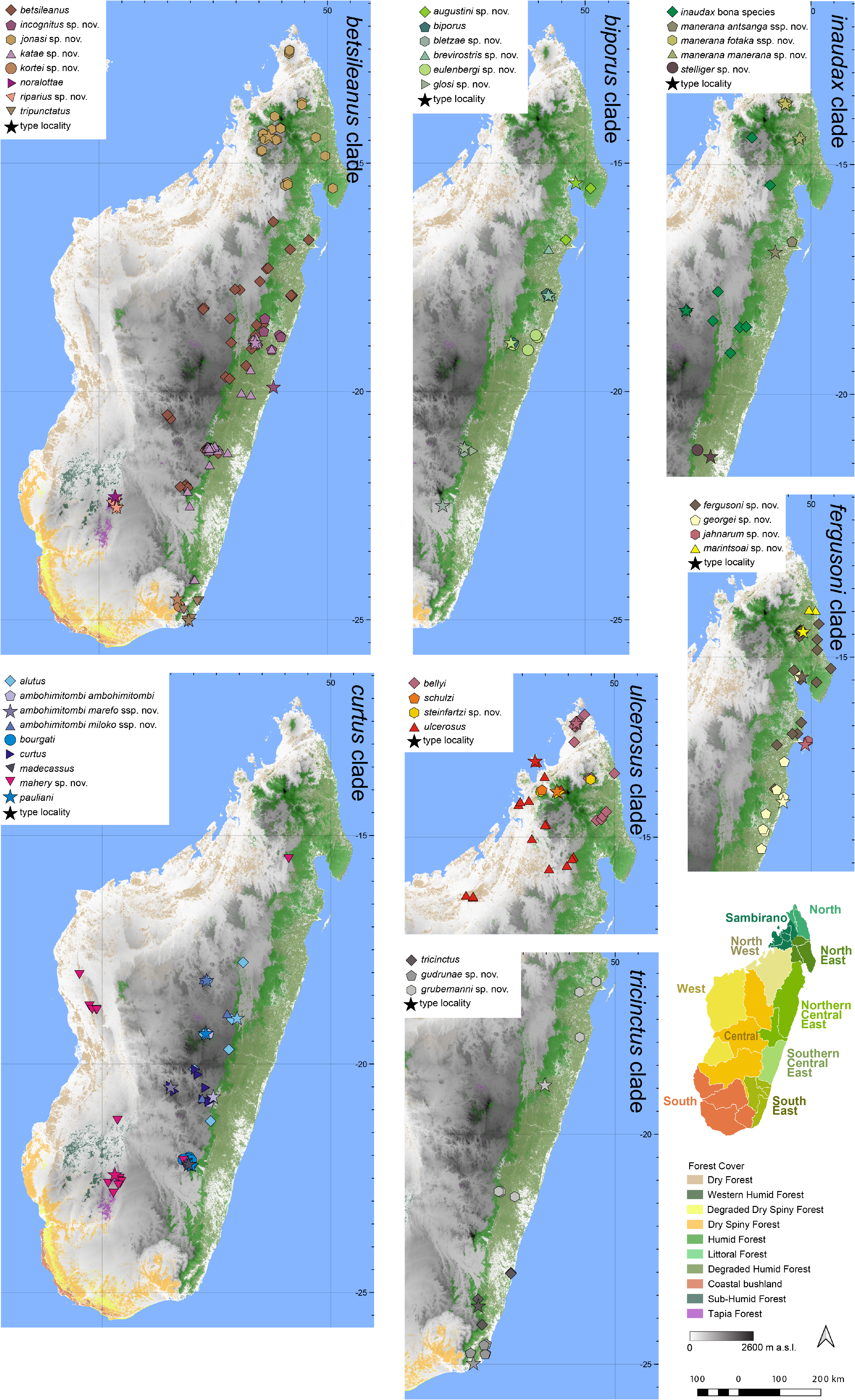
Distribution.— Endemic to the central highlands of Madagascar, north of the distribution of the nominal subspecies ( Fig. 7 View FIGURE 7 ). This subspecies is currently known from Ambohitantely, and probably also from Angavokely, and Ankazomivady. A mitochondrial haplotype corresponding to this subspecies has also been detected at Ankaratra, but due to the limited information on the sampling event and the absence of specimens (only tissue sampels were collected) this record (which might also represent mitochondrial introgression) requires confirmation. Elevation range: 1520–1735 m a.s.l.
Etymology.—The subspecies name is derived from Malagasy word miloko, meaning ‘painted’, referring to the rather distinct dorsal pattern of well-delimited dark blotches characterizing this subspecies. The subspecies name is used as a noun in apposition.


| ZSM |
Germany, Muenchen [= Munich], Zoologische Staatssammlung |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |