
Mantidactylus ambohimitombi marefo, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504344 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FF99-FF86-4F13-4ACF4FE179EE |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus ambohimitombi marefo |
| status |
subsp. nov. |
Mantidactylus ambohimitombi marefo ssp. nov.
Identity and justification.—This lineage has been considered as confirmed candidate species M. sp. 20 by Vieites et al. (2009) and M. sp. Ca20 by Perl et al. (2014). It was depicted as ‘ Mantidactylus sp. aff. pauliani “Itremo”’ by Glaw and Vences (2007). It has been collected only at its type locality, Itremo. It is characterized by rather aquatic habits and a morphology reminiscent of M. pauliani , with a short and rounded snout as in that species. Its mitochondrial DNA is near-identical to that of the nominal lineage, M. a. ambohimitombi (0.6–1.0 % pdistance) which, however, is morphologically distinct and also differs in Rag-1 haplotypes. Our phylogenomic tree confirms close relationships between this lineage and the one from Ankaratra, and for now we therefore consider a status of subspecies as adequate for the Itremo lineage, especially given that very little is known about its biology, since both its tadpoles and calls are unknown.
Diagnosis.— Mantidactylus ambohimitombi marefo is a member of the M. curtus clade, and direct sister group of M. a. ambohimitombi . See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of smooth skin, strongly expressed foot webbing with a fully webbed fifth toe, small tympanum diameter of a maximum of 8% of SVL in males, short snout, and body size of 31–36 mm distinguishes M. a. marefo from the species of the other clades in Brygoomantis ( Table 4 View TABLE 4 ). Within the M. curtus clade, distinguished from M. alutus by absence of dorsolateral ridges, absence of frenal stripe, and a shorter snout; from M. madecassus by somewhat smaller body size, less contrasted dorsal pattern and single (vs bilobed) subarticular tubercles; from M. pauliani by somewhat larger body size of males; from M. a. ambohimitombi and M. bourgati by smoother skin. Morphological distinction is most difficult from the sympatric M. curtus which appears to have a slightly lighter dorsal colour, less aquatic habits, a more pointed snout and narrower head in most specimens; these differences are obvious when comparing several individuals in the field, but not immediately apparent from the measurements taken from preserved specimens. A further difference between M. ambohimitombi marefo and M. curtus and probably all other Brygoomantis is the presence, in many individuals of this subspecies, of a bluish ring along the eye, particularly distinct posteriorly; this unique colour pattern is recognisable in life, and is not on the iris itself but on the skin surrounding the eye ( Fig. 15 View FIGURE 15 ). For detailed distinction from other new species or subspecies described herein, see the respective accounts. A full list of molecular diagnostic sites in the 16S gene of M. a. marefo in pairwise comparisons to all other Brygoomantis species (and subspecies) is provided as Supplementary appendix.
Holotype.— ZSM 762/2001 ( FGMV 2001.493 ), adult male, collected by M. Vences, D.R. Vieites, L. Raharivololoniaina, and D. Rakotomalala on 10 March 2001 in a small stream outside a forest patch at Itremo (20.6022°S, 046.5711°E, 1648 m a.s.l.), Amoron’i Mania Region, Madagascar. A 16S barcode sequence of the holotype is available from GenBank (accession AY848217 View Materials ). GoogleMaps
Paratypes.—A total of five paratypes: ZSM 761/2001 ( FGMV 2001.492 ), adult female, and ZSM 759/2001 ( FGMV 2001.476 ), ZSM 760/2001 ( FGMV 2001.478 ), ZSM 763/2001 ( FGMV 2001.495 ), three specimens of unknown sex and maturity, with the same collection data as the holotype; GoogleMaps MRSN A6747 ( FAZC 14047 ), putative female, collectd by F. Andreone and J.E. Randrianirina on 29 November 2008 in Antsirakambiaty forest GoogleMaps , Itremo.
Description of the holotype.—Adult specimen, probably a male, in mediocre state of preservation ( Fig. 9 View FIGURE 9 ). Part of right thigh muscle removed as tissue sample, and a longitudinal cut made on venter for gonad examination. Body rather slender. Head as wide as body.
Snout rounded in dorsal view; snout and head overall very short. Nostrils directed laterally, not protuberant. Nostrils nearer to tip of the snout than to eye. Canthus rostralis not clearly recognisable. Loreal region weakly concave. Tympanum recognisable, rounded, its horizontal diameter about 71% of eye diameter. Supratympanic fold present, beginning straight, and gently bending midway towards forelimb insertion. Tongue ovoid and bifid. Maxillary teeth present. Vomerine teeth not recognisable; maybe traces of vomerine teeth partly covered by tissue lateral to choanae. Choanae rounded. Subarticular tubercles single.
Inner and outer metacarpal tubercles present. Fingers without webbing. Relative length of fingers: I<II=IV<III. Finger discs slightly enlarged. Nuptial pads absent. Foot distinctly longer than tibia (114%). Lateral metatarsalia separated. Inner metatarsal tubercle present, outer metatarsal tubercle not clearly recognisable. Webbing formula: 1(0.25), 2i(1), 2e(0.25), 3i(1), 3e(0.5), 4i(1.5), 4e(1.5), 5(0.5). Relative length of toes: I<II<V=III<IV. Skin on the upper surface smooth in preservative with some scattered larger granules; in life similar. No dorsolateral folds. Ventral side smooth. Femoral glands present only as rather distinct proximal granular gland field whereas an obvious distal ulcerous macrogland appears to be missing.
Colour in preservative: dorsally uniformly brown with a light vertebral line. Weakly recognisable slightly darker crossbands on limbs. Ventrally dirty beigebrownish without clear pattern. Himdlimbs dorsally brown with light mottling. Colour in life similar to that in preservative. Dorsal side dark greyish-brown. Femoral glands light orange.
Variation.—Variation in measurements is given in Table 5 View TABLE 5 . See Fig. 15 View FIGURE 15 for colouration in life and its variation. The apparent absence of vomerine teeth in this subspecies could be of taxonomic value but is here ( Table 4 View TABLE 4 ) not included in the table of diagnostic characters as it may be variable; small aggregations of vomerine teeth recognisable in ZSM 759/2001 (in general, vomerine teeth in the M. curtus clade are present in all species except M. madecassus and M. pauliani , which Mantidactylus ambohimitombi marefo most closely resembles). In general, specimens have a characteristic appearance with a rounded and short snout, differing from most individuals of the nominal subspecies M. a. ambohimitombi , but this character is poorly reflected in the available measurements (e.g. Table 4 View TABLE 4 ), probably due to an idiosyncratic variation of various landmarks and proportions in each individual.
Possibly a slight size dimorphism, with males (30.9–34.6 mm SVL, n=4) slightly smaller than the single available female (35.9 mm SVL, n=1). Femoral glands in probable male specimens (e.g. the holotype) were rather distinctly recognisable by their light orange colour in life ( Fig. 15 View FIGURE 15 ) but not very prominent or distinct, and preserved specimens are therefore difficult to sex as seems to be typical in several taxa of the M. curtus clade.
Natural history.—Specimens were collected at Itremo in syntopy with M.curtus , a species that is morphologically similar based on standard morphometric measurements taken herein ( Table 5 View TABLE 5 ), but were immediately recognised in the field due to their distinctive habitus and aquatic habits. While M. curtus were typically found at the edge of small streams in an area of low-canopy gallery forest, and also in streams within fragments of closed forest, M. a. marefo was always found fully submerged in relatively deep pools of the stream (approximately 30–50 cm of depth), outside of dense forest.
Calls.—The call of this subspecies has not been recorded.
Tadpoles.— The tadpole of this subspecies has not been described.
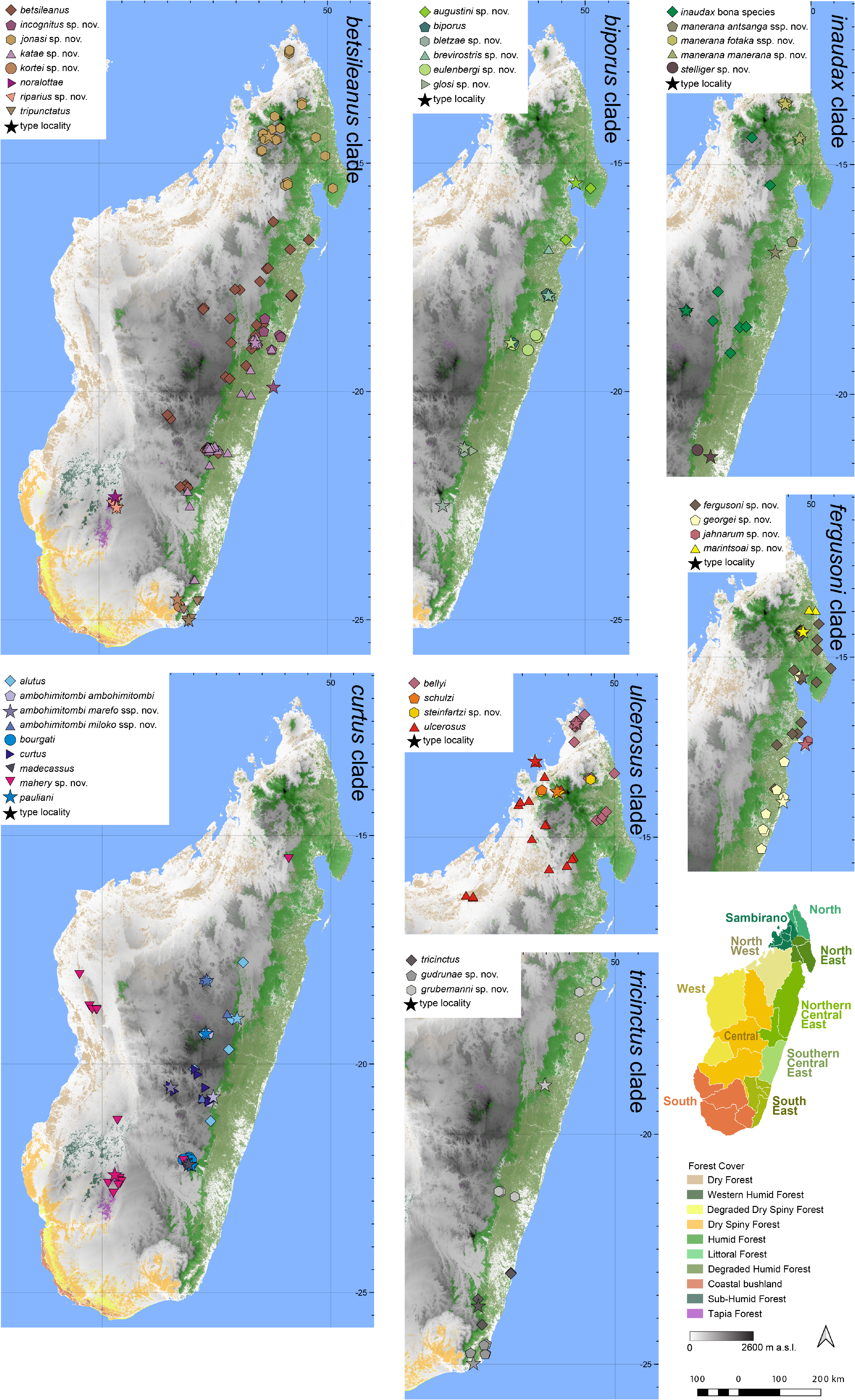
Distribution.— Apparently microendemic to the Itremo massif ( Fig. 7 View FIGURE 7 ). Elevation range: ~ 1648 m a.s.l.
Etymology.— The subspecies name is derived from the Malagasy adjective marefo, meaning ‘weak’, and refers to the surprisingly weak mitochondrial genetic divergence from its sister lineage M. a. ambohimitombi despite substantial morphological and ecological differences. The subspecies name is used as a noun in apposition.


| MRSN |
Italy, Torino, Museo Regionale di Scienze Naturali |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |