
Malthonica oceanica, Barrientos, José Antonio & Cardoso, Pedro, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.176423 |
|
DOI |
https://doi.org/10.5281/zenodo.5616030 |
|
persistent identifier |
https://treatment.plazi.org/id/03D087FC-AA70-D508-FF00-C99A1500F86C |
|
treatment provided by |
Plazi |
|
scientific name |
Malthonica oceanica |
| status |
sp. nov. |
Malthonica oceanica View in CoL sp. n.
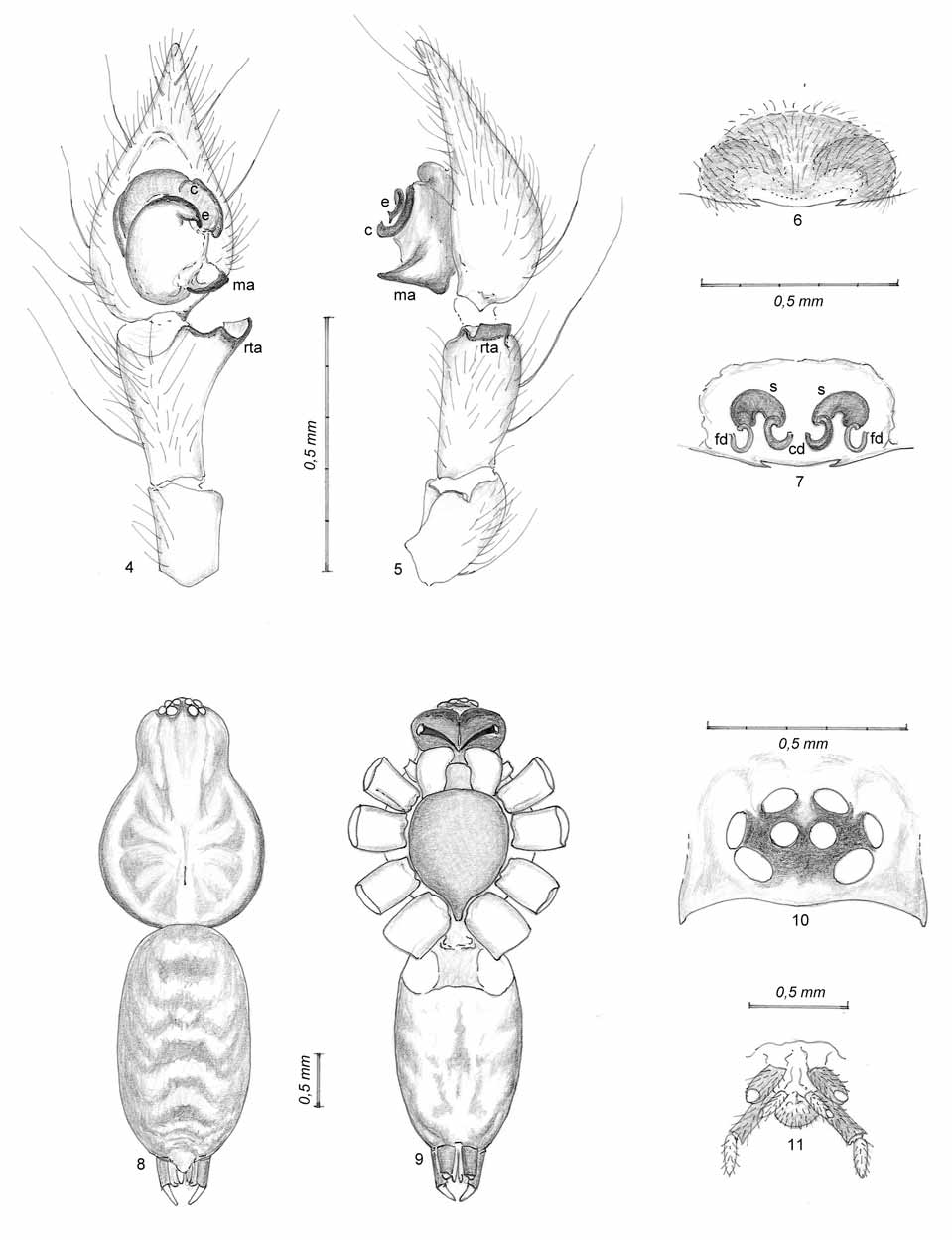
( Figs 4–11 View FIGURES 4 – 11 , 13 View FIGURES 12 – 13 , 15 View FIGURES 14 – 15 )
Type material. Type series, all from Paúl do Boquilobo Nature Reserve, Portugal (N39º 23.385 W 008º32.478): male holotype, 08.X.2002; female paratype 1, 16.VII.2002; male paratype 2, 08.X.2002; female paratype 3, 16.VII.2002; male paratype 4, 08.X.2002; male paratype 5, 08.X.2002; female paratype 6, 16.VII.2002; female paratype 7, 16.VII.2002.
The type material is deposited as follows: male holotype and female paratype 1, in Museu Nacional de Ciencias Naturales de Madrid ( Spain); paratypes 2 and 3, in the Natural History Museum of Denmark; paratypes 4 to 7 in the zoological collection of the Universidad Autónoma de Barcelona ( Spain).
Diagnosis. Comparing the two Iberian species of the genus, Malthonica lusitanica and Malthonica oceanica sp. n., a number of characters allow an easy recognition of the species. In males, while M. lusitanica presents a ventral apophysis besides the retrolateral tibial apophysis (rta) with irregular borders ( Figs 1–2 View FIGURES 1 – 3 ), the rta is the only one and has a smooth appearance in M. oceanica sp. n. ( Figs 4–5 View FIGURES 4 – 11 ). The embolus (e) is strongly developed in M. lusitanica , describing an almost complete circle, being reduced to a small appendix in M. oceanica sp. n.; the conductor (c) is much more developed in the first than in the latter species. Finally, the median apophysis (ma) is well defined in M. lusitanica with a spatulate appearance, being less defined, with a larger base and a pointed tip in M. oceanica sp. n.. In females, the development of a posterior transversal plate in M. lusitanica , the epigynal bar (eb), compared with its absence in the epigynum of M. oceanica sp. n., allows an easy recognition ( Figs 3 View FIGURES 1 – 3 , 6 View FIGURES 4 – 11 ).
Etymology. The name refers to the oceanic environment of the area where the species is found; adjective.
Description. Coloration and hair pattern of body (table 1, Figs 8–9 View FIGURES 4 – 11 ): Two irregular longitudinal dark bands in the carapace, with diffused pigmentation in a series of radial markings, a median light yellowish band with a well defined longitudinal stripe corresponding to the fovea and narrow light bands on both sides (close to the margin of the carapace, in the thoracic part). Considerably darker eye area ( Fig. 10 View FIGURES 4 – 11 ). Carapace covered by thin and sparse white hairs, together with some very thin brown hairs, especially laterally and in its posterior area. Dark reddish coloration of the chelicerae with barely visible lateral condyles and three denticles preceded by long dense hairs in the front margin, especially in the apical area; a series of five minute denticles at the rear margin is preceded by a larger tooth, apical and separated from the rest. Ventral body view presenting sparse white hairs, barely visible, and conspicuous long, straight brown or black hairs. Dark sternum, with higher density of brown pigment in the lateral areas close to the light brown coxae. Opisthosoma with straight brown hairs, transparent hairs and some feathery hairs, small and barely visible (only on the dorsal view). Brown pigmentation covers the dorsal area, creating well defined inverted v-shaped patches, especially in the posterior half. Dorsal area considerably darker than the ventral area, loosing intensity at both sides and turning into a light yellow ventral area. Spinnerets grouped in the typical disposition of the family, with very dark pigmentation, particularly the basal segments ( Fig. 11 View FIGURES 4 – 11 ).
Eye group: Typical eye formation for the genus. Both eye rows with clear procurved disposition when seen from above and clypeus narrower than the diameter of the lateral anterior eyes. Diameter of the male holotype eyes as follows (larger diameter, in mm): lateral anterior = 0.11, median anterior = 0.07, lateral posterior = 0.09, median posterior = 0.09.
Legs and pedipalps (tables 2–3): All with brown-yellow coloration, darkening distally. Ventral femora with three dark stripes, somewhat irregular and disappearing laterally. Dark patellae and a couple of annulations on tibiae, contrasting with the rest of the segment. Progressively denser leg pilosity distally. Spines also more abundant distally, except for spineless tarsi. Dorsal trichobothria on tibiae, metatarsi and tarsi. Scopulate tarsi. Paired claws pectinate in the ventral face (with five visible denticles) and unpaired claw with no visible denticles, reaching two-thirds of the other claws’ length.
Cardoso leg. carapace carapace total body basal segment of apical segment of reference length width length posterior spinneret posterior spinneret coxa trochanter femur patella tibia metatarsus tarsus total Male genitalia: Tibia widening distally, especially in the retrolateral side which presents a single apophysis – the retrolateral tibial apophysis ( Figs 4–5 View FIGURES 4 – 11 ) - wide, blunt and curved. Sides and internal face of the tibia with many hairs, including a number of long and thick hairs, especially the latter. Cymbium with numerous heterogeneous hairs, denser distally and with several longer and thicker hairs. Cymbium ending in a narrow tip. Basal part of the tegulum with a pointed projection ( Fig. 5 View FIGURES 4 – 11 ), the median apophysis, and in front of it, a slightly arched and more sclerotized structure corresponding to the embolus plus conductor. A wide tegular area, oval-shaped, occupying the two basal thirds of the alveolus ( Fig. 4 View FIGURES 4 – 11 ). Distal part of the bulb with a wide internal conduct in a semi-circle and a couple of dark curved ridges at the external side.
Female genitalia: Epigynum ( Fig. 6 View FIGURES 4 – 11 ) with a sclerotized area of semi-circular shape; at the median posterior area, close to the epigastric furrow, between two minute spine-like formations, a sub-membranous hairless area is present. Some diffused shadows denote the spermathecae. The internal ducts ( Fig. 7 View FIGURES 4 – 11 ), differentiate three symmetrically arranged paired elements: the copulatory ducts, the spermathecae and the fertilization ducts. Copulatory ducts turning 250º, from their beginning in the central section to their connection with the spermathecae. These are wide, without apparent delimited divisions, curved in an open semicircle towards the back and have a high degree of sclerotization, allowing their easy recognition. Fertilisation ducts, arising laterally from the spermathecae, being less sclerotized, forming a semicircle and becoming almost invisible close to the epigastric furrow.
Natural history
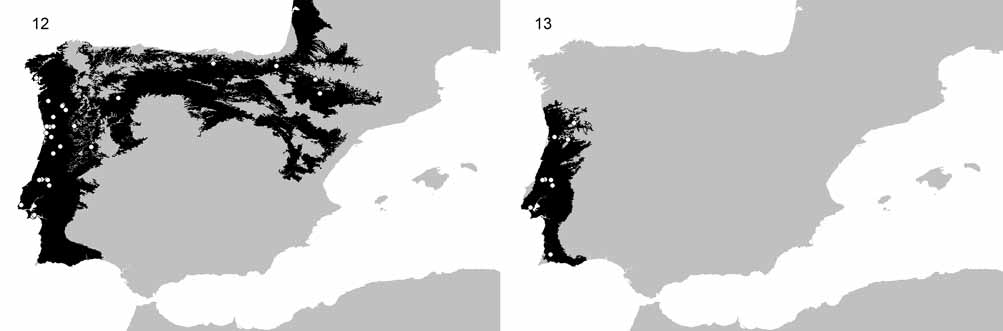
Distribution. Although both species have been described from Portugal and being this the region where they are apparently most common, Malthonica lusitanica seems to be more widespread than Malthonica oceanica sp. n.. The predicted distribution of the first species occupies almost the entire Atlantic area of the Iberian Peninsula, including the coastal areas of Portugal, northern Spain and extending towards south-western France ( Fig. 12 View FIGURES 12 – 13 ). The distribution range of the newly described species seems to be more restricted, being only known from Portugal but possibly extending its distribution towards Galicia in north-western Spain ( Fig. 13 View FIGURES 12 – 13 ). Nevertheless, where Malthonica oceanica sp. n. is present, usually M. lusitanica also occurs, often even at the same time of the year.
Phenology. The phenology of both species presents remarkable similarities, with both being mostly captured in pitfall traps during October ( Figs 14–15 View FIGURES 14 – 15 ). This abundance peak is mostly caused by the large abundance of females. On the other hand, there are also some differences.
Malthonica lusitanica presents a first spring/summer peak, which is exclusively caused by males. The species is also present during all the winter period, although in relatively low abundance. The data suggests that a single generation occurs every year or that two generations are superimposing each other.
Malthonica oceanica sp. n. presents the same first peak that in this case is caused by both males and females, during late spring and early summer. It is almost absent from the winter collecting. It is apparent then, that this species has two generations every year, each corresponding to the richness peaks of spider diversity in Mediterranean regions (Cardoso et al. in press).
Habitat types. Both species show a clear preference for sites with a dense tree cover and a relatively deep litter. It is hard therefore to predict which species occurs in which habitat type. Moreover, as already noted, they frequently live in syntopy, both spatial and temporal.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |