
Lycenchelys melanostomias Toyoshima, 1983
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4762.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:BEBD8F0D-1347-4A44-86D4-2915433D2E7B |
|
DOI |
https://doi.org/10.5281/zenodo.3809745 |
|
persistent identifier |
https://treatment.plazi.org/id/006C5E1A-FF8B-FF83-3EC6-B2B0FD35AB9D |
|
treatment provided by |
Plazi |
|
scientific name |
Lycenchelys melanostomias Toyoshima, 1983 |
| status |
|
Lycenchelys melanostomias Toyoshima, 1983 View in CoL
(Japanese name: Ohotsuku-hebigenge)
( Figs. 21–25 View FIGURE 21 View FIGURE 22 View FIGURE 23 View FIGURE 24 View FIGURE 25 ; Table 6)
Lycenchelys View in CoL sp.: Shiogaki, 1982: 23 (species list).
Lycenchelys melanostomias Toyoshima, 1983: 271 View in CoL , 333, figs. 25–27, pl. 157 (original description, type locality: southern Okhotsk Sea, Hokkaido Island, Japan); Toyoshima, 1984: 293, pl. 274-A (brief description); Toyoshima, 1985: 170, figs. 6–7, 26–27, 31, tables 1, 4 (description); Hatooka, 1993: 902, unnumbered fig. (key to species); Anderson, 1994: 113, 117 (species list); Amaoka et al., 1995: 240, pl. 402 (brief description); Koyanagi, 1997: 538, fig. 3 (brief description); Hatooka, 2000: 1032, unnumbered fig. (key to species); Hatooka, 2002: 1032, unnumbered fig. (key to species); Anderson & Fedorov, 2004: 18 (species list); Imamura et al., 2004: 84, figs. 1–3, table 1 (synonymy of Lycenchelys melanostomias and Lycenchelys brevimaxillaris ); Shiogaki et al., 2004: 71 (species list); Imamura et al., 2005: 1, figs. 1–3, table 1 (synonymy of Lycenchelys melanostomias and Lycenchelys brevimaxillaris ); Shinohara & Anderson, 2007: 64 (key to species); Kitagawa et al., 2008: 95, unnumbered fig. (brief description); Shinohara et al., 2009: 724 (species list); Amaoka et al., 2011: 315, unnumbered fig. (brief description); Balushkin et al., 2011: 1026 (species list); Hatooka, 2013: 1226, 2078, unnumbered fig. (key to species); Nakabo & Hirashima, 2015: 217 (species list and etymology of scientific name).
Lycenchelys brevimaxillaris: Toyoshima, 1985: 174 View in CoL , figs. 6–7, 29–30, 31, table 1 (original description, type locality: off Aomori Prefecture, Pacific coast of Honshu, Japan); Hatooka, 1993: 902, unnumbered fig. (key to species); Anderson, 1994: 117 (species list); Imamura, 1998: 31, fig. 9 (brief description); Hatooka, 2000: 1033, unnumbered fig. (key to species); Hatooka, 2002: 1033, unnumbered fig. (key to species); Anderson & Fedorov, 2004: 16 (species list); Imamura et al., 2004: 84, figs. 1–3, table 1 (synonymy of Lycenchelys melanostomias and Lycenchelys brevimaxillaris ); Shiogaki et al., 2004: 71 (species list); Imamura et al., 2005: 1, figs. 1–3, table 1 (synonymy of Lycenchelys melanostomias and Lycenchelys brevimaxillaris ).
Materials examined
Holotype: HUMZ 77572 , male, 182.3 mm SL, Kitami-Yamato Bank, Okhotsk Sea (44°19.5’N, 145°01’E), 915–925 m depth, 11 Oct. 1978 GoogleMaps ;
Other specimens (41 specimens, 119.3–227.6 mm SL): HUMZ 189817 (holotype of Lycenchelys brevimaxillaris ), female, 185.8 mm SL, off Aomori Prefecture, Tohoku District, northwestern Pacific (41°13’N, 141°44’E), 690–750 m depth, 18 Jan. 1982; HUMZ 152380, 152384, 152404–05, 152411, 157545, 177188, 178572–75, 182565, 192724, 192804, 4 males and 10 females, 124.0– 195.1 mm SL, Tohoku District, northwestern Pacific; HUMZ 177263, 192453, 205157, 228057, 228067–68, 228070, 228073, 228089, 5 males and 4 females, 119.3–193.2 mm SL, eastern Hokkaido Island, northwestern Pacific; HUMZ 120346, 121158, 121161, 121461, 124054–55, 124116, 126101, 126215–16, 126219–22, 126230, 126359, 126361, 10 males and 7 females, 120.6–227.6 mm SL, northeastern Hokkaido Island, Okhotsk Sea.
Diagnosis. Vertebrae 22–25 + 93–102 = 117–124; head length 11.6–15.0% SL; interorbital pore 1; occipital pores usually 2; postorbital pores usually 4; suborbital pores 6–7 + 2–3; preoperculomandibular pores usually 9; vomerine teeth 3–10; palatine teeth 2–11, usually arranged in single row (sometimes 1–2 rows); opercular flap well developed; pelvic-fin base positioned posterior to lower edge of gill opening; lateral line complete and positioned ventrally; scales absent on pectoral fin and its base; body uniformly grayish-brown when fresh.
Description. Counts and proportional measurements in Table 6.
Body very elongate, cross section oval anteriorly, compressed laterally near tail; its width at anal-fin origin 3.8–7.0 (5.0)% SL. Head short, ovoid; dorsal profile of head sloping extremely gently from posterior edge of eye to above opercular flap. Head of males slightly longer than of females in adults. Snout short, 74.4–131.0% of eye diameter (eye damaged in holotype). Eye ovoid, moderately large. Interorbital space narrow, width 12.7–33.8% of eye diameter (eye damaged in holotype). Nostril tube short, not reaching upper lip when depressed. Mouth subterminal. Posterior edge of upper jaw reaching vertical through posterior part of eye in adult males, reaching vertical through anterior margin of pupil in females and juveniles. Labial lobe of lower jaw tending to be more developed in large males than in females and juveniles (labial lobe of lower jaw damaged in holotype). Teeth on jaws sharp; upper jaw with 2–3 rows anteriorly, 1 or 1–2 rows posteriorly (1); anteriormost teeth larger than other teeth; lower jaw with 2–5 irregular rows anteriorly, 1 or 1–2 rows posteriorly (1); vomerine and palatine teeth small and conical; vomerine teeth irregularly arranged; palatine teeth usually in single row, but sometimes 1–2 rows (no data for holotype). Lower edge of gill opening slightly above lower end of pectoral-fin base. Opercular flap well developed. Gill rakers short and triangular ( Fig. 23 View FIGURE 23 ). Pseudobranch filaments short. Lateral line deciduous, complete and positioned ventrally; originating posterior to last postorbital pore and terminating on tail. Scales small and cycloid, present on body and tail, except for area around pelvic fin. Scales covering basal portions of dorsal and anal fins anteriorly; extent of scaled areas gradually increasing posteriorly, except at margins. Head, nape, pectoral axilla, pectoral fin, pectoral-fin base and area around pelvic fin without scales.
Dorsal-fin origin posterior to vertical through pectoral-fin base; 1st dorsal-fin pterygiophore between neural spines of 2nd to 5th (between 2nd and 3rd) vertebrae. Anal-fin origin below 17th to 20th (18th) dorsal-fin ray; 1st anal-fin pterygiophore posterior to parapophysis of ultimate or penultimate (penultimate) abdominal vertebra. Last dorsal-fin pterygiophore between neural spines of 2nd to 5th (between 2nd and 3rd) preural vertebrae. Last analfin pterygiophore between hemal spines of 3rd to 5th (between 3rd and 4th) preural vertebrae. Caudal fin with 1–3 (2) epural, 3–4 (4) upper hypural and 3–4 (3) lower hypural rays. Pectoral fin moderately short, not quite reaching middle of abdomen; its posterior margin rounded dorsally and having notches ventrally. Upper end of pectoral-fin base about on lateral midline of body. Pelvic fin short; its base posterior to lower edge of gill opening; its posterior margin reaching about vertical through pectoral-fin base.
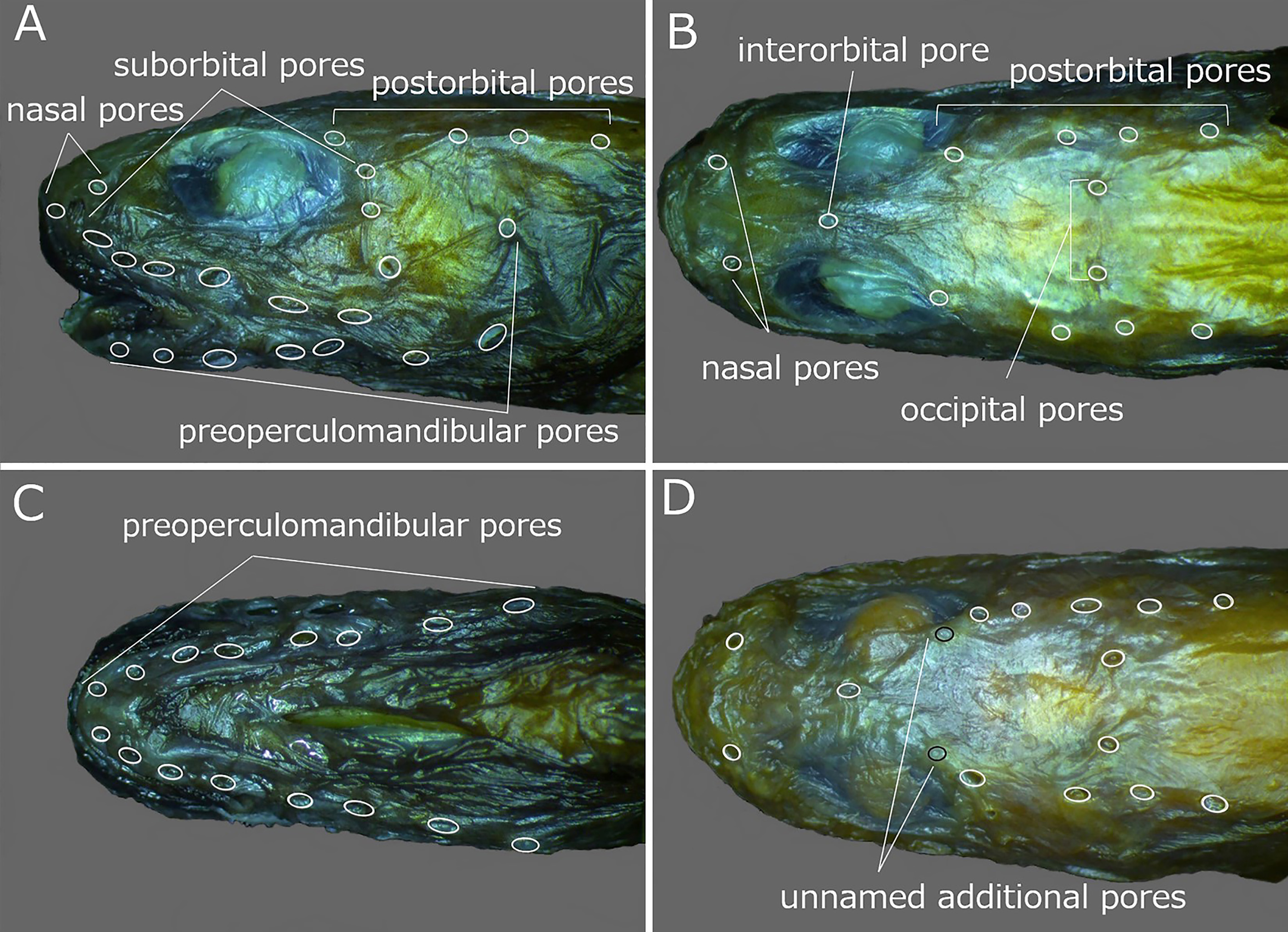
Head pores well developed and distinct. Nasal pores 2; anterior pore in front of nostril tube, posterior pore above line of 1st suborbital pore ( Fig. 22A, B View FIGURE 22 ). Postorbital pores usually 4, rarely 5 (5); when 4, distance between 1st and 2nd pores longest of those between adjacent pores; when 5, 1 additional pore present between 1st and 2nd pores ( Fig. 22A, B View FIGURE 22 ). Suborbital pores 8–10 (unknown for holotype), 6 or 7 pores located below eye and remaining 2 or 3 pores on ascending part of suborbital canal behind eye; 5th pore below posterior margin of pupil; last pore of those below eye located posterior to vertical through posterior margin of eye ( Fig. 22A View FIGURE 22 ). Preoperculomandibular pores usually 9 (9), rarely 8; 4 on lower jaw, 2 at junction of lower jaw and preopercle, and 3 on preopercle; 2 pores at junction of lower jaw and preopercle united into 1 pore in some specimens and preoperculomandibular pore series counted as 8; 2nd and 3rd pores united into 1 pore on right side of HUMZ 126361 and series counted as 8; last preoperculomandibular pore located posterior to lower part of eye ( Fig. 22A, C View FIGURE 22 ). One interorbital pore located on dorsal midline anterior to middle of eyes ( Fig. 22B View FIGURE 22 ). Occipital pores usually 2 (2), usually positioned on either side of dorsal midline; only 1 pore on right side in HUMZ 182565; occipital pore(s) located anterior to 3rd postorbital pore ( Fig. 22B View FIGURE 22 ). Two additional unnamed pores present in interorbital space on either side in line with postorbital pores, behind posterior to posterior margin of pupil only in HUMZ 178574 ( Fig. 22D View FIGURE 22 ).
Color in alcohol. Color of holotype ( Fig. 24 View FIGURE 24 ) unknown owing to head, body, and fins lacking skin. Long preserved holotype of L. brevimaxillaris with brownish head, pectoral fin and vertical fins, uniformly grayish brown body and gray abdomen. Other recently preserved specimens with dark brown head, pectoral fin and vertical fins, uniformly pale brown body and blackish abdomen.
Color when fresh (based on color photograph of HUMZ 124054; Fig. 21 View FIGURE 21 ). Head and pectoral fin black; body and vertical fins uniformly grayish brown; margin of vertical fins slightly darker and abdomen pale purplish.
Distribution. Southern Okhotsk Sea, off northwestern Pacific coast of eastern Hokkaido Island and in the northwestern Pacific from Aomori to Ibaraki prefectures, at depths of 425–1440 m ( Shiogaki, 1982; Toyoshima, 1983, 1984, 1985; Hatooka, 1993, 2000, 2002, 2013; Anderson, 1994; Amaoka et al., 1995, 2011; Koyanagi, 1997; Imamura, 1998; Anderson & Fedorov, 2004; Imamura et al., 2004, 2005; Shiogaki et al., 2004; Shinohara & Anderson, 2007; Kitagawa et al., 2008; Shinohara et al., 2009; this study).
Size. The largest specimen examined during this study measured 227.6 mm SL (230.8 mm TL), about equal to the previously recorded maximum length of 23 cm TL ( Hatooka, 2013).
Remarks. Lycenchelys melanostomias is similar to L. hippopotamus , L. makushok and L. rassi in having more than 100 total vertebrae, 1 interorbital pore, 1–2 occipital pores, 4–5 postorbital pores, a single lateral line positioned ventrally and no distinct spots or blotches on the body (vs. lacking this combination of characters in other species of Lycenchelys ) (e.g., Toyoshima, 1983, 1985; Fedorov & Andriashev, 1993; Anderson, 1995; Imamura et al., 2004; Shinohara & Anderson, 2007; this study). See Remarks under accounts of L. hippopotamus , L. makushok and L. rassi for detailed comparisons of L. melanostomias with each.
Two additional pores behind the posterior margin of the pupil were found in 1 specimen of L. melanostomias ( Fig. 22D View FIGURE 22 ). These pores have not been previously described for L. melanostomias , although similar pores are known in Lycenchelys parini Fedorov, 1995 ( Fig. 25 View FIGURE 25 ). Lycenchelys melanostomias also resembles L. parini in having similar counts, proportional measurements and arrangements of head pores (e.g., dorsal-fin rays 111–120 vs. 118, and anal-fin rays 98–107 vs. 105, head length 11.6–15.0 vs. 12.2% SL, postorbital pores usually 4 vs. 4 and suborbital pores 8–10 vs. 9 in L. melanostomias respectively; Table 6) ( Fedorov, 1995a; Imamura et al., 2004, 2005; this study). However, L. melanostomias is easily separable from L. parini in having the lateral line positioned ventrally rather than midlaterally on the side ( Fedorov, 1995a).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lycenchelys melanostomias Toyoshima, 1983
| Kawarada, Shumpei, Imamura, Hisashi, Narimatsu, Yoji & Shinohara, Gento 2020 |
Lycenchelys brevimaxillaris: Toyoshima, 1985: 174
| Imamura, H. & Shinohara, G. & Shiogaki, M. 2005: 1 |
| Anderson, M. E. & Fedorov, V. V. 2004: 16 |
| Imamura, H. & Machida, Y. & Ohta, S. 2004: 84 |
| Shiogaki, M. & Ishito, Y. & Nomura, Y. & Sugimoto, T. 2004: 71 |
| Hatooka, K. 2002: 1033 |
| Hatooka, K. 2000: 1033 |
| Imamura, H. 1998: 31 |
| Anderson, M. E. 1994: 117 |
| Hatooka, K. 1993: 902 |
| Toyoshima, M. 1985: 174 |
Lycenchelys melanostomias
| Nakabo, T. & Hirashima, Y. 2015: 217 |
| Hatooka, K. 2013: 1226 |
| Amaoka, K. & Nakaya, K. & Yabe, M. 2011: 315 |
| Balushkin, A. V. & Sheiko, B. A. & Fedorov, V. V. 2011: 1026 |
| Shinohara, G. & Narimatsu, Y. & Hattori, T. & Ito, M. & Takata, Y. & Matsuura, K. 2009: 724 |
| Kitagawa, D. & Imamura, H. & Goto, T. & Ishito, Y. & Fujiwara, K. & Ueda, Y. 2008: 95 |
| Shinohara, G. & Anderson, M. E. 2007: 64 |
| Imamura, H. & Shinohara, G. & Shiogaki, M. 2005: 1 |
| Anderson, M. E. & Fedorov, V. V. 2004: 18 |
| Imamura, H. & Machida, Y. & Ohta, S. 2004: 84 |
| Shiogaki, M. & Ishito, Y. & Nomura, Y. & Sugimoto, T. 2004: 71 |
| Hatooka, K. 2002: 1032 |
| Hatooka, K. 2000: 1032 |
| Koyanagi, M. 1997: 538 |
| Amaoka, K. & Nakaya, K. & Yabe, M. 1995: 240 |
| Anderson, M. E. 1994: 113 |
| Hatooka, K. 1993: 902 |
| Toyoshima, M. 1985: 170 |
| Toyoshima, M. 1984: 293 |
| Toyoshima, M. 1983: 271 |
Lycenchelys
| Shiogaki, M. 1982: 23 |