
Lithoxus raso, Silva & Covain & Oliveira & Roxo, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4347.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:DD5FC393-AFE1-4DEF-8261-FA54663E2B06 |
|
DOI |
https://doi.org/10.5281/zenodo.6047707 |
|
persistent identifier |
https://treatment.plazi.org/id/434FDD1E-CD51-6425-339C-4EA1F977F977 |
|
treatment provided by |
Plazi |
|
scientific name |
Lithoxus raso |
| status |
sp. nov. |
Lithoxus raso , sp. n.
Fig. 5 View FIGURE 5 ; Table 1
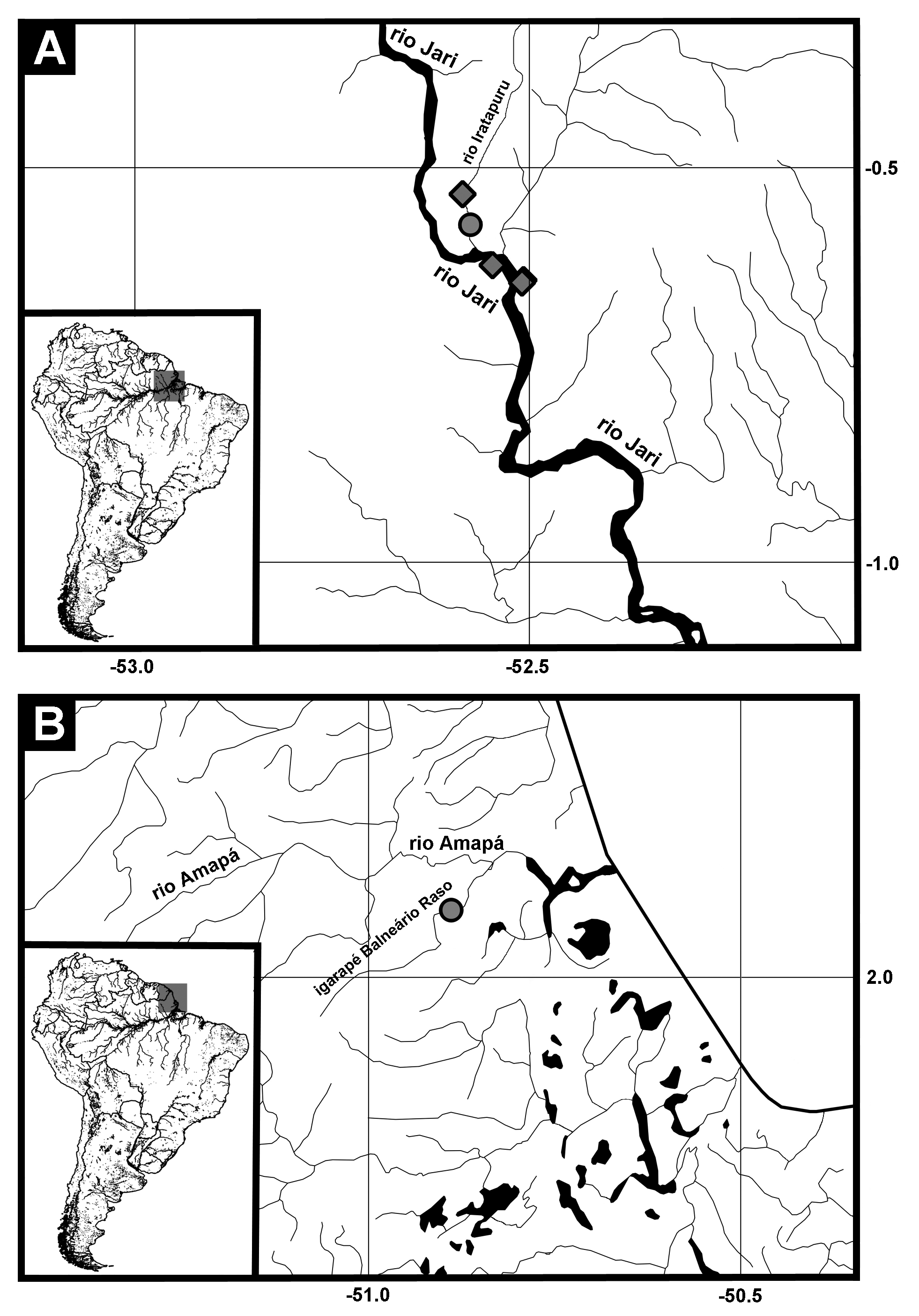
Holotype. MZUSP 123132, 44.3 mm SL, Brazil, municipality of Amapá, Amapá States, igarapé Balneário Raso, rio Amapá basin, 02°05’25” N, 50°53’20” W, 0 2 December 2015, C. Oliveira & B.F. Melo.
Paratypes. LBP 21184 (5, 28.8–42.9 mm SL), collected with holotype . MHNG 2767.052 View Materials (2, 25.3–44.8 mm SL), collected with holotype .
Diagnosis. Lithoxus raso differs from all congeners, except L. pallidimaculatus by the presence of large light spots all over the body, spots similar in size with orbit diameter, ( Fig. 5 View FIGURE 5 ; vs. absence of light spots over the body or light spots very small, similar to size of a tooth, covering only the dorsal portion of the head in L. boujardi ; see Fig. 6 View FIGURE 6 for coloration pattern among Lithoxus species); from L. surinamensis and L. pallidimaculatus by having an adipose fin (vs. adipose fin absent); from L. lithoides and L. jantjae by having five branched anal-fin rays (vs. four); and from L. jantjae by having 14 branched caudal-fin rays (vs. 12). Moreover, the new species differs from L. bovallii by having a lower premaxillary tooth cup length, 3.3–4.0% HL (vs. 4.2–10.4% HL, personal communication with J.W. Armbruster for measurements of L. bovallii specimens from the type locality); from L. boujardi by having a greater adipose–spine length, 7.2–8.1% SL (vs. 3.6–4.7% SL), and a greater dorsal adipose– caudal distance, 13.1–14.6% SL (vs. 10.3–11.7% SL); from L. planquettei by smaller caudal peduncle depth, 8.4– 8.6% SL (vs. 9.7–10.5% SL); and from L. stocki by having medial premaxillary teeth with an enlarged and widened cusp (vs. cusps of all premaxillary teeth similar in size, not enlarged), and by having a shorter cleithral width, 30.9– 31.2% SL (vs. 32.6–34.2% SL).
Description. Morphometric data presented in Table 1. Medium sized loricariid (42–46 mm SL). In lateral view, dorsal profile convex from snout tip to dorsal-fin origin; straight and gradually descending from dorsal-fin origin to adipose-fin origin; concave from adipose-fin origin to upper caudal-fin spine. Ventral profile slightly concave from snout tip to anal-fin origin; slightly convex from anal-fin origin to insertion of lower caudal-fin spine.
Head and snout broadly rounded. Snout elongated (50–54% HL), anterior region depressed. Area around tip of snout free of plates and odontodes. Posterior nostril small, half of eye diameter. Dorsal profile of head ascending approximately 45° to parieto-supraoccipital. Eye small (orbit diameter 13–15 % HL), dorsolaterally positioned. Superior margin of orbit not elevated. Mouth moderate in size; oral disk occupying almost entire ventral surface of head. Lips without odontodes; lower lip larger than upper lip, but far from reaching cleithrum region. Margin of oral disk fringed, with low triangular papillae. Maxillary barbel relatively long and projecting anterolaterally from upper lip. Teeth bicuspid with deep division between cusps; four left premaxillary teeth, six to eleven left dentary teeth. Premaxillary tooth cusps increasing in size from lateralmost to medialmost tooth with medialmost tooth cusp two times as large as lateralmost tooth cusp. Dentary tooth cusps similar in size to lateralmost premaxillary tooth cusps. Dentary teeth of same size.
Body dorsoventrally depressed. In dorsal view, greatest body depth at dorsal-fin origin. Greatest body width at cleithral region; body progressively narrowing from opercular region to caudal fin. Cross-section of body between pectoral and pelvic fins rounded dorsally and flat ventrally. Body almost entirely covered by plates; except in ventral portions of head, abdomen and along dorsal-fin base.
Dorsal surface of body entirely covered by plates; three predorsal plates; seven plates below dorsal-fin base; four plates between dorsal fin and adipose fin. Lateral surface of body entirely covered by plates; mid-dorsal plate series developed, reaching end of adipose fin; lateral median plates 23–25; mid-ventral plate series developed, reaching end of adipose fin. Body plates with minute odontodes. Odontodes slightly hypertrophied on pectoral-fin spines, becoming gradually larger distally. Longest odontodes on posteriormost evertible cheek plates.
Dorsal-fin II,7; dorsal-fin spinelet V -shaped, dorsal-fin locking mechanism present; dorsal-fin origin approximately at midpoint between pectoral- and pelvic-fin origins; last dorsal-fin ray not reaching adipose fin when depressed. Pectoral-fin I,6; pectoral-fin spine reaching slightly beyond base of pelvic-fin spine when depressed. Pelvic-fin I,5; reaching anal-fin origin when depressed. Anal-fin I,5. Adipose fin with single azygous preadipose plate; posterior margin of adipose-fin membrane convex, extending posteriorly beyond adipose-fin spine. Cross-section of caudal peduncle ellipsoid, rounded dorsally and ventrally. Three caudal peduncle plate rows. Caudal-fin I, 7−I,7; caudal fin obliquely forked, lower lobe longer than upper.
Color in alcohol. Background color dark brown on dorsum and sides of body, becoming lighter ventrally. Large white spots on trunk, becoming larger on posterior portion of body. Fins with irregular and poorly defined bars: three or four on anal, pectoral, dorsal and pelvic-fin rays. Caudal-fin dark with one large hyaline bar in middle portion. Hypertrophied odontodes on posteriormost evertible cheek plates and on pectoral-fin spines; pectoral-fin spines reddish. Cross-section of caudal peduncle ellipsoid.
Sexual dimorphism. Males possess a papilla posterior to urogenital opening (absent in females).
Etymology. The specific epithet raso is in reference to the type-locality, the igarapé Balneário Raso, a tributary of the rio Amapá, Atlantic coastal drainage. A noun in apposition.
Distribution. Lithoxus raso is only known from igarapé Raso, at the municipality of Amapá, in Amapá State, Brazil ( Fig. 3 View FIGURE 3 ).
Habitat. The new species L. raso is found associated with rocks and sand in the bottom of small to medium sized rivers ( Fig. 4 View FIGURE 4 ).
! " # $%& $ ’
()! * + +, + .
! " # # # $ %
$%& /0// 121 13 144 5./16.3 7 L. planquettei 8 9 + 8:!;6;6; </=11/0=
64/ <4.////
$%&>.46<5.?146.0 7 L. planquettei $:!;8; 6; &!
$%& /0/3 1<0 .1 4=. 5.44/6.0 7 L. planquettei 8:!;6;6; <14 64/ 0
$%& /0/3 1<0 .1 4=3 5.4436.0 7 L. planquettei 8:!;6;6; <14 64/ 0
$%& /0// 121 13 140 5.41<6.0 7 L. planquettei 8 9 + 8:!;6;6; </=11/0=
64/ <4.////
$%& /2=/ 14. 12 3 /= 5.4/.6.0 7 L. planquettei 8 9 8:!;6;6; </=12220
64/ <4.3==?
$%& /2=/ 104 12 <1= 5.4/36.0 7 L. planquettei 8 9 & @ 5:!;6;6; <</====? 64/ 33=/4
$%& ?? 144 5.0 0 0 6.0 7 L. planquettei 8 9 A B,:!;6;6; <4<432..
64/.4.?<<<
$%& ?? 14= 5.00=6.0 7 L. planquettei 8 9 A B,:!;6;6; <4<432..
64/.4.?<<<
$%& ?? 14? 5.00?6.0 7 L. planquettei 8 9 A B,:!;6;6; <4<432..
64/.4.?<<<
$%& /2=/ 103 12 3=<5.4//6.0 7 L. planquettei 8 9 C 5:!;6;6; <<0304 64/ </=?0// /1<2= =.1/0 7 $?2/=?. L. jariensis D D; E F;C; @ 61 4332 64/ 4=<0 $%& /0/<110 1. 3.= 5 //<06.0 7 L. 8 9 B:!;6; H; 4 ..?....
pallidimaculatus 644 /=3.220
$%& /0/<112 1. 31. 5 //<26.0 7 L. 8 9 B:!; 6; H; 4 ..?....
pallidimaculatus 644 /=3.220
$%& /203 141 14.=? 5 //4/6.0 7 L. pallidimaculatus 8 E 8 H::!;6;,; 3?10?0//
644 400////
$%& /0// 1== 1=./// 5 //=16.0 7 L. 8 H $ 8 H 8, E:!; 6 4 /01 =333
pallidimaculatus ;8, E; 64<?/14442
. $%& /0// 1== 1=.//3 5 //=.6.0 7 L. 8 H $ 8 H 8, E:!; 6 4 /01 =333
pallidimaculatus ;8, E; 64<?/14442
$%& /0// 1== 1=.//<5 //=/6.0 7 L. 8 H $ 8 H 8, E:!; 6 4 /01 =333
pallidimaculatus ;8, E; 64<?/14442
$%& /0// 1== 1=.//1 5 //0?6.0 7 L. 8 H $ 8 H 8, E:!; 6 4 /01 =333
pallidimaculatus ;8, E; 64<?/14442 ……continued on the next page I8 J
! " # # # $ %
$%& /0/<124 K1</0<5.?1?6.0 7 L. bovallii B H 8 " 9:!;6;:; <24??.20
64= 2021442
$%& /0</ 134 10.4<5 //216.0 7 L. bovallii>:!,:!;6 / <...3=?
;,; 642?/<0///
$%& /032 1<2 ./ 5.=1?6.0 7 L. bovallii $ $ B:!;6;,; <<//04 640.?.000=
1?<I.J
$%& 1. <13 5 //<=6.0 7 L. bovallii H $ & H:!;6;6; &!
$%& /040 102 .<.1=3 5.=<16.0 7 L. bovallii $ 8 H & H:!;6;,; <3?4<<<< 6 44 0==?<<<
$%& /032 131 ./ <40 5.0?46.0 7 L. bovallii $ $ B:!;6;,; <<//04 640.?.000=
$%& /032 131 ./ <4= 5.0?26.0 7 L. bovallii $ $ B:!;6;,; <<//04 640.?.000=
$%& /032 1<2 ./ 5.=.16.0 7 L. bovallii $ $ B:!;6;,; <<//04 640.?.000=
1?<I/J
$%& /0// 142 1. </= 5 //<?6.0 7 L. bovallii H $ & H:!;6;6; &!
$%& /0// 142 1. </? 5 //416.0 7 L. bovallii H $ & H:!;6;6; &!
$%& /040 102 .<.1=<5.=<.6.0 7 L. bovallii $ 8 H & H:!;6;,; <3?4<<<<
644 0==?<<<
$%& /032 1<2 ./ 5.=..6.0 7 L. bovallii $ $ B:!;6;,; <<//04 640.?.000=
1?<I3 J
$%& .4 /3? 5.2<06.0 7 L. stocki *:!;6;6; 3 3? 643?/4=
$%& .4 <4. 5.0./6.0 7 L. stocki *:!;6;6; 3 3?.? 643 =3..
$%& .4 /<1 5.2<=6.0 7 L. stocki *:!;6;6; 3 3? 643?/4=
$%& /0/3 1<. .1 432 5.4416.0 7 L. boujardi 8 8 H C:!;6;6; <1=1/<64/ 2=<=.
$%& D5.36?I.J 5.<?<6.0 7 L. boujardi * C:!;6;6; <13333 64/ 4.20
$%& D5.36. 5.<=36.0 7 L. boujardi & 8 8 H C:!;6;6; &!
$%& /22/ 1?1 13.4<5.4.16 .0 7 L. boujardi C L 8 H C:!;6;6; <11<2220
64/ 2.33==?
$%& /22/ 1?1 13.4= 5.4..6.0 7 L. boujardi C L 8 H C:!;6;6; <11<2220
64/ 2.33==?
$%& D5.36?I/J 5.<?46.0 7 L. boujardi * C:!;6;6; <13333 64/ 4.20
$%& /01= 1?2 $ <44 5 /./46.0 7 L. boujardi & 8 H C:!;6;6; &!
$%& /0/3 1<. .1 431 5.4<?6.0 7 L. boujardi 8 8 H C:!;6;6; <1=1/<64/ 2=<=. 41/4 /4?2<7 01.0/2 H. ancistroides : 68;: 8;; @ &!



| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |