
Leiopathes annosa, Wagner, Daniel & Opresko, Dennis M., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3974.2.11 |
|
publication LSID |
lsid:zoobank.org:pub:6141ACE0-3131-48B0-9348-05AF452D1A17 |
|
DOI |
https://doi.org/10.5281/zenodo.6093526 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA8C77-9C2B-FFAA-FF58-FAC1FA0FFEA7 |
|
treatment provided by |
Plazi |
|
scientific name |
Leiopathes annosa |
| status |
sp. nov. |
Leiopathes annosa View in CoL sp. nov.
( Fig. 1 View FIGURE 1 a–e, 2a–d, 3a–h, 4a–o, 5a, 6a–d)
not Leiopathes glaberrima, (Esper 1972) View in CoL (and later authors; see Opresko and Baron-Szabo, 2001 for synonymy) Leiopathes glaberrima, Grigg and Opresko, 1977: 242 View in CoL –261, fig. 7. Leiopathes glaberrima, Grigg 1988: 142 View in CoL .
Leiopathes glaberrima, Chave and Malahoff, 1998: 40 View in CoL , fig. 93. Leiopathes glaberrima, Roark et al., 2006 View in CoL *: 1–14.
Leiopathes glaberrima, Parrish and Baco, 2007 View in CoL *: 186.
Leiopathes View in CoL sp. Roark et al. 2009 *: 5204–5208.
Leiopathes View in CoL sp. Wagner et al., 2011: 211–225, fig. 1k, 2d.
Leiopathes View in CoL sp. Wagner et al., 2012: 76, 80, 84, 88, 90.
Leiopathes View in CoL sp. P4-227-2 Brugler et al., 2013: 325, 327, 335, fig. 2–3. Leiopathes View in CoL USNM1070976 Brugler et al., 2013: 335, fig. 2–3.
* Re-identification based on in situ photographs in HURL archives only. For all others, both specimens and in situ photographs were examined.
Type locality. South of Ni‘ihau Island, Main Hawaiian Islands, 21°49’21”N 160°6’37”W, 382 m.
TABLE 1. Leiopathes specimens examined as part of this study (* = type material; NR = not reported; 1 molecular sequence data for specimen BPBM D1871 (as P4-227-2) was reported in Brugler et al. (2013) and is available from GenBank under accession numbers KF054599 View Materials , KF054665 View Materials and KF054466 View Materials ; 2 molecular sequence data for specimen USNM 1070976 was reported in Brugler et al. (2013) and is available from GenBank under accession numbers KF054593 View Materials , KF054663 View Materials and KF054467 View Materials ).
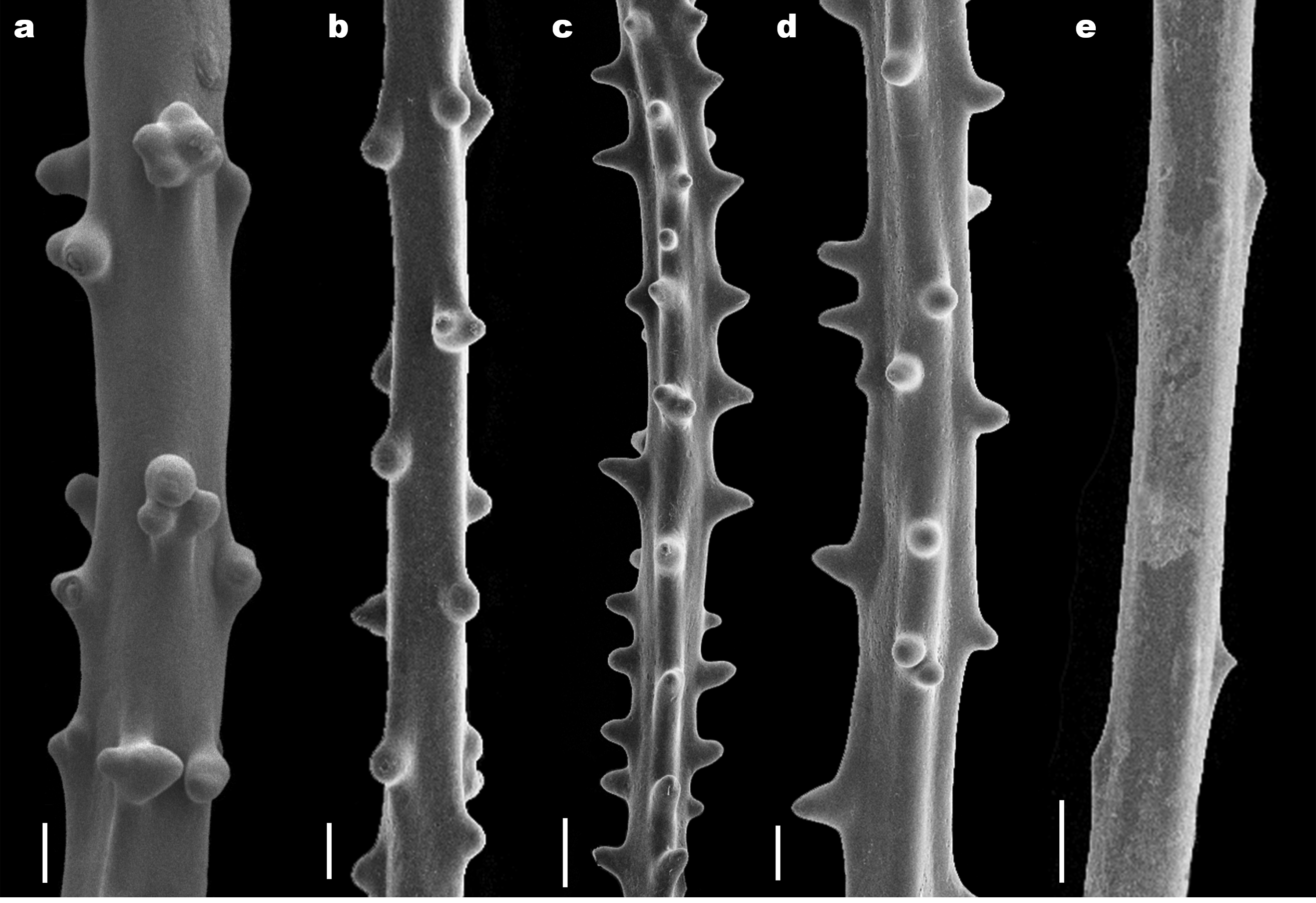
Diagnosis. Colonies are up to 1 m in height or more, with most branches located in an approximately single plane like a fan ( Fig. 1 View FIGURE 1 a–c, 3a–h). In some colonies, branches overlap considerably producing a rather thick corallum that is still planar, whereas in others all the branches are in a single plane ( Fig. 1 View FIGURE 1 a–c, 3 a–h). Most of the smaller branches are directed distally relative to the branches from which they arise, as well as upwards towards the top of the corallum. Branching is mostly irregularly bilateral, but uniserial in some places, with the branches tending to be directed vertically. The highest order branches are typically 1.5–6 cm in length, 1.4 mm in diameter at their midpoint with tissue (range = 1.0– 1.8 mm), and on average spaced 0.8 cm apart (range = 0.4–1.7 cm). Polyps are variable in size (0.88–3.35 mm), arranged irregularly on all sides of the corallum on thicker branches and uniserially on terminal branches with adjacent polyps spaced 1.2–3.2 mm apart, resulting in 5–6 polyps/cm ( Fig. 1 View FIGURE 1 ). Spines are typically 75 Μm tall (range = 30–225 Μm) and 140 Μm wide at the base (range = 35–315 Μm). On the terminal branchlets (~ 0.07 mm in diameter excluding the spines), the spines can be hemispherical, balloon-shaped, irregularly knobby or with one or more round or misshapen lobes ( Fig. 2 View FIGURE 2 d, 4a–o). On such terminal branchlets, 3–5 spine rows are visible in lateral view ( Fig 2 View FIGURE 2 d, 4a–o). On thicker branches (0.15 mm in diameter excluding spines), the spines are more hemispherical to mound-like with fewer, but larger round lobes, and the spine arrangement in rows becomes obscured ( Fig. 2 View FIGURE 2 b–c). On branches thicker than 0.27 mm in diameter excluding the spines, the spines become smaller and more knob-like or somewhat conical to deltoid in shape, with little remaining evidence of lobes ( Fig 2 View FIGURE 2 a). The living tissues are colored bright orange ( Fig. 1 View FIGURE 1 a, 3a–h).
Material examined. Holotype ( BPBM D1871), south of Niʻihau Island, Main Hawaiian Islands, 21°49’21”N 160°6’37”W, 382 m, HURL submersible Pisces IV, dive P4-227, specimen 2, 2 December 2009, ethanol. Paratype ( USNM 77480), bank north of Necker Island, Northwestern Hawaiian Islands, no coordinates recorded, 457 m, R/ V Townsend Cromwell, cruise 76-06-73, fish trawl, 15 October 1976, dry. For all other non-type material examined as part of this study see Table 1.
Description of holotype. The holotype (BPBM D1871) consists of two separate ethanol-preserved fragments collected from the distal part of a colony ( Fig. 1 View FIGURE 1 a–b). The colony from which the holotype fragments were collected, branches sparsely towards the center of the corallum, and becomes more dense and irregular distally, with multiple orders of branching ( Fig. 1 View FIGURE 1 a–b, 3a). The first holotype fragment has a basal branch diameter of 5 mm and is approximately 31 cm long and 16 cm wide, whereas the second fragment has a basal branch diameter of 6 mm, measures 33 cm in length and 18 cm in width. On both fragments polyps are arranged irregularly on all sides of the axis on the main branch and larger branches, and uniserially on the terminal branches. Polyps are 0.88–3.35 mm in transverse diameter, and spaced 1.2–3.2 mm apart on the terminal branches, resulting in 5–6 polyps/cm. Terminal branches are typically 3–4 cm in length (range = 1.5–6 cm), 1.4 mm in width at their midpoint with tissue (range = 1.0– 1.8 mm) and 280 Μm in width without tissue (range = 165–400 Μm). On the terminal branches, 3–4 spine rows are visible in lateral view; however, on thicker branches the spine arrangement in rows becomes obscured ( Fig. 2 View FIGURE 2 a–b). Spines are smooth and typically hemispherical, but spines with multiple lobes occur often ( Fig. 2 View FIGURE 2 c–d). On terminal branches spines are generally 70 Μm tall (range =30–100 Μm) and 145 Μm wide (range=70–310 Μm). On thicker branches (width = 400–810 Μm without tissue), spines range between 30–110 Μm in height and 130–315 Μm in width ( Fig. 2 View FIGURE 2 a).
Description of paratype. The paratype (USNM 77480) consists of a dried colony that is 63 cm tall, 17 cm wide, 8 mm in basal diameter and was broken off above the base ( Fig. 1 View FIGURE 1 c, e). The branching is sparse towards the center of the corallum, and becomes more dense and irregular distally, with nine orders of branching ( Fig. 1 View FIGURE 1 c). Most of the smaller branches are directed distally relative to the branches from which they arise, as well as upwards towards the top of the corallum ( Fig. 1 View FIGURE 1 c, e). Branching is mostly irregularly bilateral, but uniserial in some places, with the branches tending to be directed vertically ( Fig. 1 View FIGURE 1 c, e). Branches coming off the main axis range between 3–5 mm in diameter. The terminal branches are 1.5–6 cm long, tend to be curved distally, and measure 400–650 Μm at their midpoint with remaining tissue. On portions without remaining tissue, terminal branches measure 280–440 Μm at their midpoint. Despite being preserved as a dry specimen, polyp remnants remain on a few terminal branchlets of the paratype ( Fig. 1 View FIGURE 1 e) and measure 2–2.5 mm in diameter. On the terminal branches, 4–5 spine rows are visible in lateral view; however, on thicker branches the spine arrangement in rows becomes obscured. Spines on terminal branches are smooth and range between hemispherical, balloon-shaped and irregularly knobby. On terminal branches (250–400 Μm at their midpoint without spines), spines are 55–140 Μm tall and 75–240 Μm wide at their base. On thicker branches (width = 450–675 Μm without tissue), spines range between 50–120 Μm in height, 120–260 Μm in width and tend to be mound-like, conical or deltoid in shape.
Intraspecific variability and a unique morphotype. The holotype and paratype of Leiopathes annosa sp. nov. both have skeletal spines that are smooth, hemispherical, often times multi-lobed, 30–225 Μm in height and 35–310 Μm in width. In the examination of the 14 additional specimens assigned to this species (see Table 1), skeletal spine morphology ranged widely even within the same specimen, and varied from hemispherical, balloonshaped, irregularly knobby, multi-lobed, mound-like, conical and deltoid ( Fig. 4 View FIGURE 4 a–o). However, spines with multiple lobes were present on all specimens assigned to L. annosa sp. nov. (see Table 1). Similar to skeletal spine morphology, the dimensions of skeletal spines varied between the 16 specimens assigned to L. annosa sp. nov. (Table 1), and ranged from an average height of 55 Μm and average basal width of 125 Μm for specimen USNM 1071417, to an average height of 95 Μm and average basal width of 190 Μm for specimen USNM 99500. However, the skeletal spines of all specimens assigned to L. annosa sp. nov., ranged between 30–225 Μm in height and 35– 310 Μm in width at the base. In contrast, one specimen from Hawaiian waters ( Leiopathes sp.; USNM 52278) was unique in that its skeletal spines were (1) much smaller (average = 40 Μm; range = 20–65 Μm), (2) narrower (average = 90 Μm; range = 30–160 Μm), and (3) had almost no multi-lobed spines ( Fig. 4 View FIGURE 4 p). Additionally, while the polyp spacing of specimen USNM 52278 (1.2–3.05 mm) is similar to those of the specimens assigned to L. annosa sp. nov (1.2–3.2 mm), its polyps are smaller (0.70–2.10 mm vs. 0.88–3.35 mm). Furthermore, the 16 Hawaiian specimens assigned to L. annosa sp. nov. were all collected at depths ranging between 295–536 m, whereas specimen USNM 52278 was collected in much deeper waters (966 m; Table 1). Based on these substantial differences, specimen USNM 52278 represents a distinct species that is morphologically more similar to L. montana and L. glaberrima . However, a more detailed study, ideally using both morphological and molecular markers, will be needed to determine the taxonomic identity and affinities of specimen USNM 52278.
Comparisons. In Hawaiian waters, Leiopathes annosa sp. nov. has previously been referred to as Leiopathes sp. ( Roark et al. 2009; Wagner et al. 2011; Wagner et al. 2012; Brugler et al. 2013) and L. glaberrima ( Grigg and Opresko 1977; Grigg 1988; Chave and Malahoff 1988; Roark et al. 2006; Parrish and Baco 2007). As part of this study, we reexamined in situ photographs and specimens referenced in those previous studies, and as a result reassigned those records to L. annosa sp. nov. The previous misidentification of the Hawaiian specimens as L. glaberrima was due to: (1) Hawaiian specimens were not compared to specimens from other geographic locations, and (2) diagnostic characters of L. glaberrima were largely unknown because Esper's (1792) original species description was quite brief and type material remained without further reexamination until recently ( Opresko & Baron-Szabo 2001). The recent redescription of L. glaberrima has established the diagnostic characters of this species which include (1) triangular, simple and smooth spines which are 20–60 Μm tall, (2) polyps that are up to 1 mm in diameter, (3) terminal branches with a length of 1–2 cm, and (4) an irregularly branched corallum ( Opresko & Baron-Szabo 2001). Leiopathes annosa sp. nov. differs from L. glaberrima in that its spines are hemispherical and frequently multi-lobed, and substantially larger (30–225 Μm) ( Fig. 5 View FIGURE 5 a. e). Additionally, polyps of L. annosa sp. nov. reach a larger maximum size (3.35 mm) than those in L. glaberrima (1 mm), and its terminal branches are longer (<6 cm). Finally, L. annosa sp. nov. colonies are fan-like and bright orange in situ ( Fig. 1 View FIGURE 1 a, 3a–h), whereas living colonies of L. glaberrima are grayish-white, white, pale orange or bright orange, and branched irregularly ( Bo 2008; Vertino et al. 2010; Mastrototaro et al. 2010; Bo et al. 2015). Leiopathes glaberrima has been reported to occur at depths as shallow as 37 m in the Gulf of Mexico (Opresko 2009) and as deep as 1532 m off Morocco ( Grasshoff 1989), whereas L. annosa sp. nov. has a limited bathymetric range (295–536 m).
Leiopathes annosa sp. nov. can be easily differentiated from all but one of the remaining seven described species of Leiopathes on the basis of the morphology of its skeletal spines ( Fig. 5 View FIGURE 5 a–e). In L. expansa , L. grimaldi , L. valdiviae , L. secunda , L. acanthophora and L. montana , the spines are predominantly conical, subdeltoid or relatively tall (height greater than width at the base) with a rounded apex. In none of these seven species are the spines hemispherical or multi-lobed (see summary of species characteristics as given on Table 1 in Molodtsova 2011). Hemispherical spines are, however, found in L. bullosa ( Fig. 5 View FIGURE 5 b, 6f–h). The major morphological difference between L. annosa sp. nov. and L. bullosa is the fact that in the former the spines, especially those on the terminal branches, can be multi-lobed ( Fig. 4 View FIGURE 4 a–o, 6c–d), whereas in L. bullosa multi-lobed spines have not been reported ( Fig. 6 View FIGURE 6 f–h). Other differences between the two species exist in the maximum thickness of the terminal branches (0.2 mm in L. bullosa and 0.4 mm in L. annosa sp. nov.), maximum length of the terminal branches (2 cm in L. bullosa and 6 cm in L. annosa sp. nov.), maximum size of the spines (140 Μm in L. bullosa and 225 Μm in L. annosa sp. nov.), arrangement of the spines (clearly in rows in L. bullosa , and not in discernable rows in L. annosa sp. nov.), maximum size of the polyps (2 mm in L. bullosa and 3.35 mm in L. annosa sp. nov.), and in the density of the polyps (3–5/cm in L. bullosa and 5–6/cm in L. annosa ). L. annosa sp. nov. forms colonies that are large, flabellate, with relatively thick branches ( Fig. 1 View FIGURE 1 a, 3a–h). In comparison, the branches of L. bullosa are much thinner, and even though individual clusters of branches of the holotype were described as being somewhat planar, the holotype consists of small fragments, and therefore the size, shape, and branching pattern of intact colonies of L. bullosa are unknown.
Previous studies in Hawaiʻi. Several studies have reported on different ecological aspects of L. annosa sp. nov. in Hawai‘i (identified at the time as L. glaberrima or Leiopathes sp.), including on its: (1) growth rate and longevity (Roark et al. 2006; Roark et al. 2009), (2) population density ( Grigg 1988), (3) associated fish assemblages ( Parrish 2006), and (4) sexual reproduction (Wagner et al. 2011). Roark et al. (2006, 2009) used highresolution skeletal radiocarbon measurements and estimated average radial growth rates of 5–13 µm/year for this species, corresponding to longevities ranging between 350–4,265 years, thus making this the longest living marine organism studied to date. Furthermore, Roark et al. (2009) concluded that growth rates are not uniform throughout the lifespan of this species, with faster initial growth rates that gradually decrease with age. Grigg (1988) examined the population densities of various dominant deep-water corals (365–400 m) off Oʻahu Island, and noted average densities of 0.002–0.003 colonies/m2 for this species. Parrish (2006) studied fish assemblages within deep-water corals (300–500 m) across various Hawaiian Islands, and noted that various fish occasionally use taller colonies of L. annosa sp. nov. as shelter. Wagner et al. (2011) used histological techniques to study the reproductive anatomy of eight Hawaiian antipatharian species, including a sample of the L. annosa sp. nov. holotype, and noted that it had both previtellogenic and vitellogenic oocytes within its primary mesenteries. Finally, as part of a study on the evolutionary history of the order Antipatharia, Brugler et al. (2013) provided mitochondrial and nuclear DNA sequence data for two specimens (USNM 1070976 and BPBM D1871 as P4-227-2) that are assigned to L. annosa sp. nov. here (see Table 1 for GenBank accession numbers). Molecular sequences of these two specimens had identical sequences in the intergenic region between nad5 and nad1, and between cox3-cox1, but did differ in a single position in the intergenic region between trnW and nad2 ( Brugler et al. 2013). Future studies will need to include a molecular component in order to determine whether the morphological analysis presented in this study holds up, as well as to elucidate the taxonomic relationships among the various species within the genus Leiopathes .
Etymology. From the latin ‘ annosa ’ (long-lived) in reference to the extreme longevity estimates reported for this species (350–4,265 years; Roark et al. 2006; Roark et al. 2009).
Distribution. Under the name L. glaberrima or Leiopathes sp., L. annosa sp. nov., has previously been reported throughout the Hawaiian Islands ( Grigg & Opresko 1977; Chave & Malahoff 1998; Parrish and Baco 2007; Roark et al. 2006; Roark et al. 2009; Wagner et al. 2011; Wagner et al. 2012; Brugler et al. 2013), as well as from Johnston Atoll ( Chave & Malahoff 1998) at depths between 300– 500 m. Specimens examined as part of this study were collected throughout the Hawaiian Archipelago from Hawai‘i Island (19°30'00''N 155°00'00''W) to Salmon Bank (27°01'10''N 176°31'33''W) at depths between 295–536 m (Table 1).
Leiopathidae View in CoL biogeography. Within the monogeneric family Leiopathidae View in CoL , seven species have been recorded from a limited geographic region, including (1) L. expansa View in CoL from Madeira ( Johnson 1899), (2) L. montana View in CoL from the Great Meteor Seamount ( Molodtsova, 2011), (3) L. grimaldii View in CoL from two locations in the northeastern Atlantic ( Roule 1905), (4) L. valdiviae View in CoL from the Nicobar Islands ( Pax 1915; Pax 1922), (5) L. secunda View in CoL from Tasmania and New Zealand ( Opresko 1998; Cairns et al. 2009), (6) L. acanthophora View in CoL from South Australia and New Zealand ( Opresko 1998; Cairns et al. 2009), and (7) L. bullosa View in CoL from South Australia and New Zealand ( Opresko 1998; Cairns et al. 2009), ( Fig. 7 View FIGURE 7 ). In contrast, L. glaberrima View in CoL has been recorded from widespread geographic locations, including from throughout the Mediterranean ( Gray 1857; Opresko & Baron-Szabo 2001; Bo 2008; Carlier et al. 2009; Sinninger & Pawlowski 2009; Deidun et al. 2010; Vertino et al. 2010; Mastrototaro et al. 2010; D’Onghia et al. 2012; Brugler et al. 2013; Angeletti et al. 2014; Bo et al. 2014a, 2014b, 2015; Fabri et al. 2014; Ingrassia et al. in press), the northeastern Atlantic ( Johnson 1899; Grasshoff 1985, 1989; Brito & Ocaña 2004), the northwestern Atlantic and Caribbean ( Brook 1889; Cairns et al. 1993; Williams et al. 2006, 2007; Opresko 2009) ( Fig. 7 View FIGURE 7 ). To the eight previously described species of Leiopathes View in CoL , we here add L. annosa View in CoL sp. nov. from the Hawaiian Archipelago and Johnston Atoll ( Fig. 7 View FIGURE 7 ). In addition to the described Leiopathes View in CoL fauna, several undescribed and unidentified Leiopathes View in CoL spp. have been reported from around the globe, including from the coasts off Chile ( Häussermann & Försterra 2007; Yañez et al. 2009), the southeastern United States ( Fernholm & Quattrini 2008; Ross & Quattrini 2009; Brugler et al. 2013), eastern Canada ( Murillo et al. 2010), Ireland ( Roberts et al. 2006; Wienberg et al. 2008; Le Guilloux et al. 2010), Saint Paul Island ( Guerriero et al. 1988), Azores ( Carreiro-Silva et al. 2013), New Zealand ( Cairns et al. 2009) and Hawai‘i ( Parrish & Baco 2007; Baco 2007; Brugler et al. 2013; this report) ( Fig. View FIGURE 7
7). Based on all available records, the Leiopathidae View in CoL are currently known from tropical latitudes around the Nicobar Islands (7.80°N; Pax 1915; Pax 1922) to temperate latitudes off the coasts of Ireland (56.50°N; Wienberg et al. 2008) and New Zealand (48.80°S; Opresko 1998). The conspicuous absence of this family from polar regions ( Fig. 7 View FIGURE 7 ) may suggest that the geographic distribution of the Leiopathidae View in CoL is limited by oceanographic or environmental conditions. Alternatively, the absence of Leiopathidae View in CoL records from polar regions may be due to lower sampling efforts in these areas, and future collections in these regions will be needed to determine whether this is the case.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Leiopathes annosa
| Wagner, Daniel & Opresko, Dennis M. 2015 |
Leiopathes
| Brugler 2013: 325 |
| Brugler 2013: 335 |
Leiopathes
| Wagner 2012: 76 |
Leiopathes glaberrima
| Chave 1998: 40 |
Leiopathes glaberrima,
| Grigg 1988: 142 |
| Grigg 1977: 242 |