
Latrodectus umbukwane, Wright & Wright & Sole & Lyle & Tippett & Sholto-Douglas & Verburgt & Engelbrecht, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4700.4.12 |
|
publication LSID |
lsid:zoobank.org:pub:1602CD7C-66ED-4C77-86ED-40A594187F99 |
|
persistent identifier |
https://treatment.plazi.org/id/E0791125-CC25-F97A-88D3-F9CAFC13FD95 |
|
treatment provided by |
Plazi |
|
scientific name |
Latrodectus umbukwane |
| status |
sp. nov. |
Latrodectus umbukwane View in CoL new species Wright, Wright, Lyle and Engelbrecht
Zoobank ID: urn:lsid:zoobank.org:act:AAEDE7DF-8B9D-4853-8BC6-D58E61CD13C0
Type material: Holotype: ♀ ( NCA2018 /991), SOUTH AFRICA : KwaZulu-Natal: Umkhanyakude Municipality: Phinda Private Game Reserve , (27°46’26.40”S 32°20’9.46”E; 64 m. a.s.l.), sand forest, 2017-11-01, B. Wright, from tree hollow, by hand; dissected GoogleMaps . Paratypes: 2 ♂ ( NCA2018 /990), same data as holotype, captive raised GoogleMaps ; 1 ♀ ( NMSA29324 About NMSA ), same data as holotype, dissected GoogleMaps ; 3 ♂ ( NMSA29325 About NMSA ), same data as holotype, captive raised GoogleMaps ; 1 ♀ ( SAMC10028 View Materials ), same data as holotype except Lulubush, Phinda Private Game Reserve (27°47’35.9”S 32°20’33.0”E; 68 m. a.s.l.), not dissected GoogleMaps ; 3 ♂ ( SAMC10029 View Materials ), same data as holotype, captive raised GoogleMaps .
Etymology: The species epithet is derived from the isiZulu word “ umbukwane ”, meaning something eye-catching or spectacular that one cannot simply walk past it due to its profound beauty. This is in reference to the impressive size and remarkable coloration of this species. It is a noun in apposition and therefore gender neutral.
Measurements: See Tables 1 View TABLE 1 and 2 View TABLE 2 .
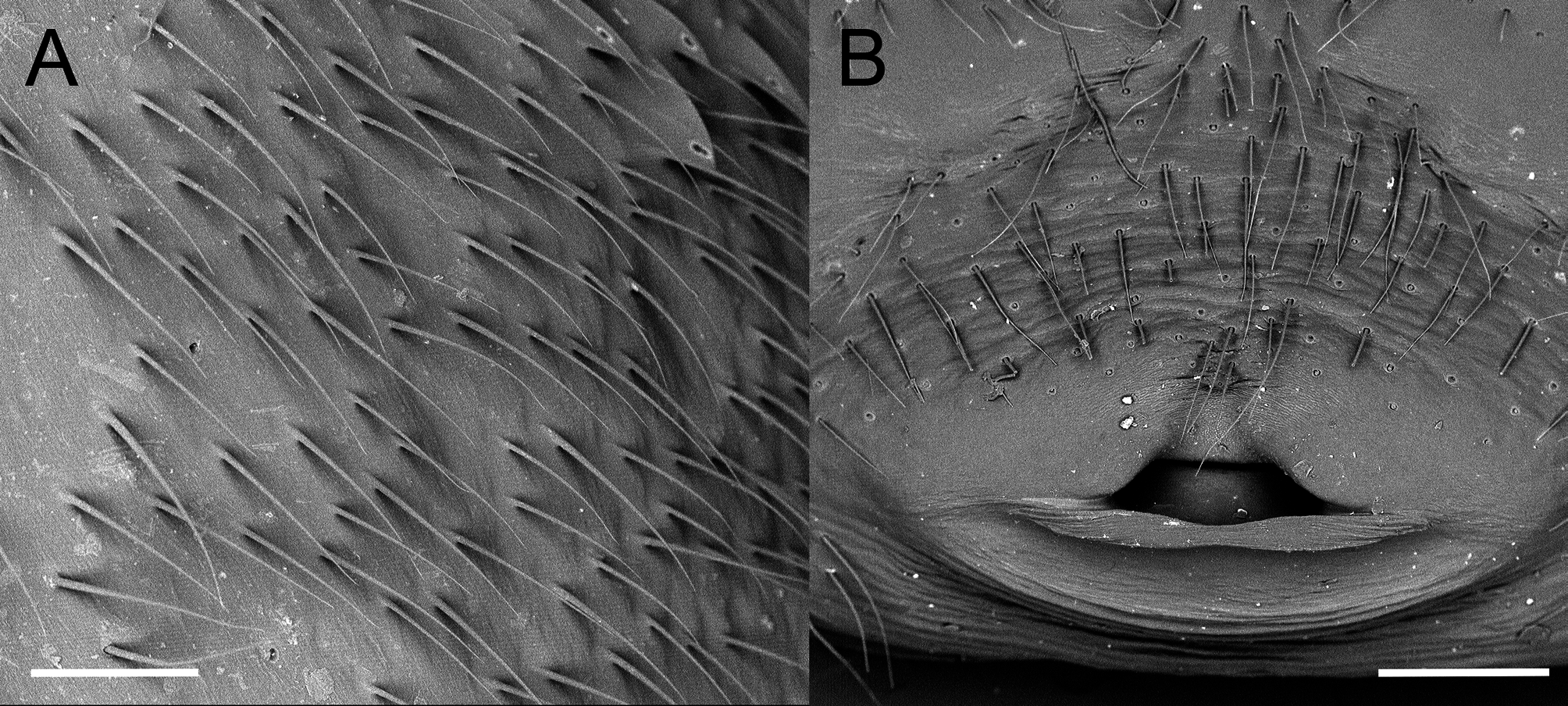
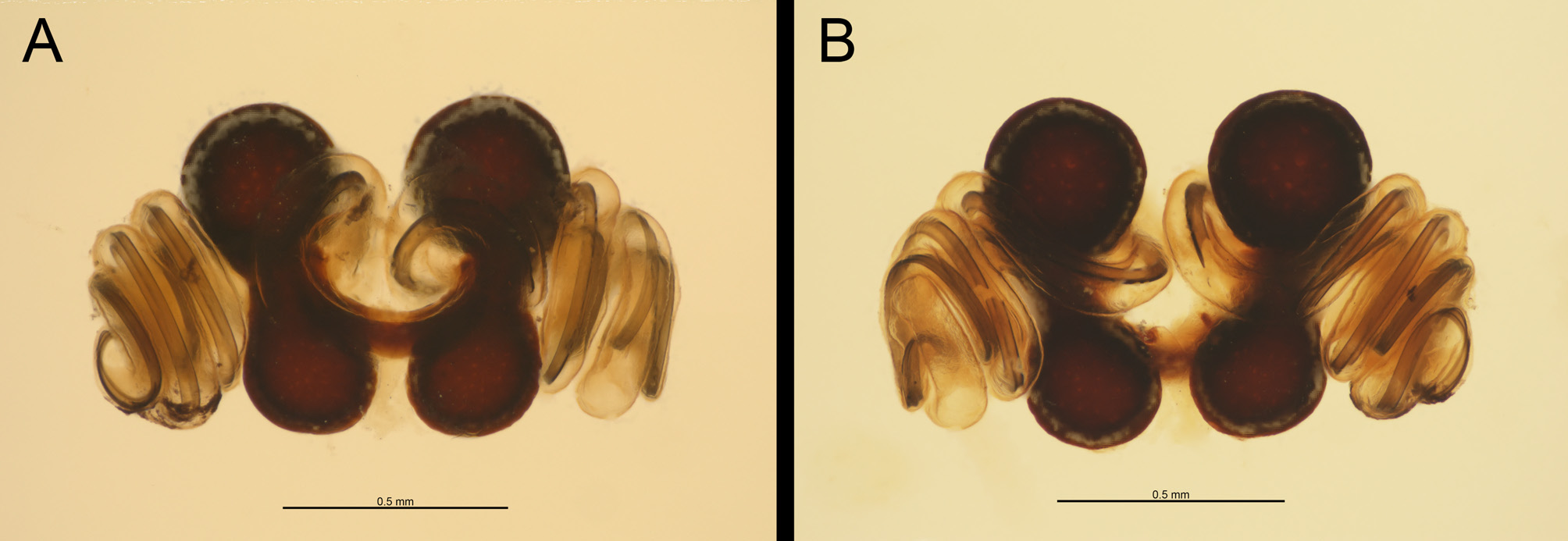
Diagnosis: Females of L. umbukwane n. sp. can be separated from African congeners by the combination of a distinct red marking on the ventral surface of the abdomen and a red stripe on the posterior dorsal surface of the abdomen ( Figs. 1 View FIGURE 1 A– C, 3A, B). Females can further be separated from other African species, except for L. geometricus and L. rhodesiensis , by having the spermathecae parallel to one another ( Fig. 5 View FIGURE 5 , 6A, B View FIGURE 6 ). They can be distinguished from L. geometricus and L. rhodesiensis by the following combination of characters: presence of thick curved abdominal setae of approximately the same length as opposed to finer setae of two different lengths in L. geometricus and L. rhodesiensis ( Fig. 4A View FIGURE 4 ; Levy 1998); ventral red marking irregular in shape and situated more posteriorly than the orange-red hourglass of L. geometricus and L. rhodesiensis (Fig. 3B); legs uniformly black in adults and not banded (Fig. 3A, B); abdomen black with light-coloured, irregular oval lines laterally extending onto the dorsal surface with a single dorsal posterior red stripe, positioned medially (Fig. 3A, B); similar pale markings, but more circular in shape, are present along the dorso-median surface of the abdomen; larger size ( Table 1 View TABLE 1 ). Females can be further distinguished from L. rhodesiensis by having three loops of the copulatory ducts as opposed to five ( Fig. 5 View FIGURE 5 , 6A, B View FIGURE 6 ).
Males of L. umbukwane n. sp. are generally similar to or slightly smaller than other African species in size ( Table 2 View TABLE 2 ). They can be diagnosed from all African congeners by having a yellow-white ventral marking anterior to the spinnerets and a yellow-white transverse ventral marking near the book lungs (Fig. 3D). Males can be distinguished from L. rhodesiensis by the four loops of the embolus as opposed to five ( Fig. 6 View FIGURE 6 C–F).
Females of L. umbukwane n. sp. can be distinguished from all other species worldwide, except for L. apicalis Butler 1877 , L. erythromelas Schmidt & Klaas 1991 , and L. obscurior Dahl 1902 , by having the spermathecae parallel to one another as opposed to a being in a V-shape ( Fig. 5 View FIGURE 5 , 6A, B View FIGURE 6 ; McCrone & Levi 1964; Kaston 1970; Abalos 1980; Levy 1998; Melic 2000; Knoflach & van Harten 2002; Berendonck & Greven 2005; Vink et al. 2008; Aguilera et al. 2009; Kananbala et al. 2010; Saaristo 2010; Goyal & Malik 2017). Latrodectus apicalis and L. erythromelas have little published data and no information was found on the shape of their spermathecae. However, L. erythromelas can be distinguished from L. umbukwane n. sp. by the small size of the adult female (total length 5.5mm-9.0mm). L. apicalis can be separated by its geographical distribution as it is considered endemic to the Galapagos. L. obscurior from Madagascar has parallel spermathecae as in L. umbukwane n. sp. but it is described as being uniformly black in colour with a red ventral marking as in L. geometricus , and was previously considered a synonym of this species ( Dahl 1902; Levi 1959; Schmidt et al. 1994). It is also described as being peridomestic, as is L. geometricus , whereas L. umbukwane n. sp. appears to be limited to pristine sand forest habitats. Therefore L. umbukwane n. sp. can be distinguished from L. obscurior on differences in coloration and habitat selection.
Behavioral traits can be used as additional diagnostic features. L. umbukwane n. sp. produces a large egg sac, approximately 27 mm long and 17 mm wide, that is bright purple when first laid ( Fig. 1C View FIGURE 1 ), changing to shiny grey over time. This differs from the smaller, spiked egg sacs of L. geometricus , the large, whitish, woolly egg sacs of L. rhodesiensis , and the smooth white egg sacs of other African species ( Lotz 1994). Fertile egg sacs are generally tear-dropped shaped (n=13), while infertile egg sacs remain rounded (n=3). The webs and selection of retreat sites also appear to be unique amongst African species.
Description: Females: Measurements in Table 1 View TABLE 1 . Carapace moderately setose on cephalic and lateral parts of thoracic region, less so on posterior part of thoracic region; fovea transverse, procured. Eyes widely separated, all similar in size; PME directly posterior to AME; PLE wider than ALE. Sternum subtriangular. Distal portion of maxillae pallid. Pedipalps with single toothed claw. Legs uniformly moderately setose throughout, tapering distally; metatarsi I and IV gently concave in lateral profile, other segments straight; three tarsal claws present, superior claws with one row of teeth; ventrally situated paired accessory claws present on all legs; tarsus IV with distinct prolateral row of spines. Abdomen moderately setose with setae all of uniform length. Epigynum variable but oval, elevated and sclerotized with continuous longitudinal procurved opening. Spermathecae dumbbell-shaped, linked by short thick duct; spermatheca 2 larger than spermatheca 1; copulatory openings situated medially; copulatory ducts thinned-walled, weakly sclerotized, coiled around duct between spermathecae and forming three-and-half loops retrolateral to spermathecal ( Fig. 5 View FIGURE 5 , 6A, B View FIGURE 6 ). Coloration of adult in life uniformly black with distinctive red marking ventrally on abdomen situated between spiracles and spinnerets; vertical red stripe present on posterior dorsal surface extending from spinnerets, fading proximally; elongated, irregular white loops on lateral to dorsolateral surface of abdomen; similar but more circular markings are median dorsal surface. Males: Measurements in Table 2 View TABLE 2 . Carapace sparsely setose with long setae; fovea transverse, procurved. Eyes proportionally larger relative to carapace than in female, eye pattern as in female except PER closer to AER. Maxillae uniform in colour. Mating bulbs with embolus having four to five coils ( Fig. 6 View FIGURE 6 C–F), located distally on cymbium, last coil extending medially, curving downwards along retrolateral edge of palp ( Fig. 6D View FIGURE 6 ), continuing ventrally, curving upwards and terminating in a long thin point; embolus originating retrolaterally, near rounded cymbial apophysis ( Fig. 6E View FIGURE 6 ); median tegular apophysis sharply bent, almost 90° degrees, ending in blunt point ( Fig. 6E View FIGURE 6 ). Legs moderately setose with short setae; tarsi lacking accessory claws. Abdomen sparsely covered in elongated setae. Coloration medium to dark brown, cephalothorax paler ventrally. Legs pale brown, darker towards joints and proximal half of femur I dark. Abdomen with white marking ventrally anterior to spinnerets, white longitudinal marking on posterior dorsal surface starting at spinnerets and fading proximally; lateral surfaces with elongated irregular white markings. All white markings with margins darker than the rest of the abdomen. In older specimens these white markings may be absent.
Habitat and conservation status: All known localities for the new species fall within sand forest habitats ( Fig. 2 View FIGURE 2 ). Sand forest is a distinct lowland forest type that is dry, and occurs only on flat terrain ( Fig. 1D View FIGURE 1 ). It is limited to KwaZulu-Natal province and neighboring parts of Mozambique, and is considered a critically endangered vegetation type ( Mucina & Geldenhuys 2006). Of the seven wild female specimens recorded, six were found in pristine, tall sand forest with predominantly bare ground below the canopy, on Phinda Private Game Reserve, which is part of the larger Munyawana Conservancy. A single specimen was found in the garden of the manager’s house on Tembe Elephant Park, at the edge of disturbed sand forest. The potential distribution of L. umbukwane n. sp. is shown in Figure 2 View FIGURE 2 under the assumption that it is limited to sand forest habitats. The AOO is ca. 698 km 2 and the EOO is 4963 km 2. Twenty-three percent of the AOO falls within nationally protected areas in South Africa and Mozambique, and a substantial additional proportion of sand forest is protected in privately protected areas. Assuming the potential distribution developed ( Fig. 2 View FIGURE 2 ) is sufficiently accurate, the extinction risk status for L. umbukwane n. sp. would be considered as Least Concern ( IUCN 2012). However, given the uncertainty surrounding the ecology of this species, we recommend that a status of Data Deficient is assigned. Additional surveys of nearby coastal forests are required to confirm the assumption that this species is limited to sand forest, but coastal forests have been surveyed for spiders in the past by both professional and citizen arachnologists, and we are not aware of the species having been located during such surveys. Surveys of sand forest should also be a priority to confirm the true extent of occurrence of this species. Climatic conditions across the predicted geographic range of the species are a mean maximum temperature of 32–33.1°C, mean minimum temperature of 10.7–11.6°C, and an annual rainfall of 660–818 mm.
The selection of retreat sites also appears to be unique in this species. All specimens located to date were at an elevated position of more than 50 cm above the ground, with a silk retreat in a tree hollow. Webs were not generally present outside the hollow, but when they were recorded, they were typical hackle webs radiating outward from the hollow, and can be extensive ( Fig. 1E View FIGURE 1 ). Unlike webs of other Latrodectus which are closer to the ground with many strands of silk attached to the ground surface, the webs of L. umbukwane n. sp. are largely anchored to higher branches, with only a few strands of silk going to ground in a few cases. No sticky silk threads were observed being connected to the ground in the wild or captivity.
Development: Spiderlings ( Fig. 7A View FIGURE 7 ) typically emerge from the egg sac 29.9 days after laying (27–36 days; n=9). We assume that emerging spiderlings are in the second instar as observed in other Latrodectus species (Foster & Kingford 1983). Spiderlings were observed actively moving in some egg sacs for a day or two before emerging, possibly waiting for optimum conditions. Egg sacs produce an average of 584 spiderlings (471–692; n=4). General abdominal shapes, patterns, and positions remain relatively constant in both sexes throughout life, changing only in colour and clarity. Spiderling coloration is golden brown with white dorsal median and lateral abdominal markings on emergence ( Fig. 7A View FIGURE 7 ). These markings are relatively constant in shape and position for the duration of life, but change in colour and prominence. In third instars the dorsal median and lateral markings remain white but are more prominent ( Fig. 7B View FIGURE 7 ). In the fourth instar the dorsal median and lateral markings are greenish-white with indistinct dark brown outlines ( Fig. 7C View FIGURE 7 ). In later instars the dorsal median and lateral markings turn yellow with white outlines, resembling small fried eggs ( Fig. 7D View FIGURE 7 ). Overall coloration, including the legs which are banded in immatures, darkens until black at maturity after ca. 6 months.
In immature males, the palpal tarsus becomes swollen ca. 15–20 days after emergence. They reach maturity at ca. 26 days. Mature male abdominal coloration fades dramatically as the spider ages. The colour of the abdomen of a young mature male is brown with light yellow-white oblong blotches dorso-medially and laterally, with indistinct dark brown outlines, and no transverse light areas (Fig. 3A). Old mature males have a dark brown abdomen with darker brown irregular circle shapes only visible under bright light or with flash photography.
Phylogenetic relationships: COI sequencing yielded a total of 422 nucleotide base pairs. The identification function on BoLD did not match the COI barcodes to any specimen or species in the database, despite BoLD including sequences for 14 species of Latrodectus . The BLAST search on GenBank provided a weak identity match of 87% with a number of genera including Steatoda Sundevall 1833 , Crustulina Menge 1868 and several species of Latrodectus . The ML tree is presented with bootstrap support (BS) and posterior probabilities (PP) overlaid ( Fig. 7 View FIGURE 7 ). Two lineages, labelled I and II correspond to the tredecimguttatus species-group and the geometricus speciesgroup respectively, are supported. L. umbukwane n. sp. falls into lineage II. L. umbukwane n. sp. appears as a wellsupported lineage (BS: 100 %; PP: 1.00), sister to L. geometricus (BS: 54 PP: 0.57). The relationship between L. umbukwane n. sp. and L. geometricus is corroborated by morphological evidence as the two species share parallel spermathecae. The mean inter-specific pairwise genetic distances between. L. umbukwane n. sp. — L. geometricus is 14.7%. Several of the species in lineage I are para- or polyphyletic.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |