
Lanice kellyslateri, Lavesque & Daffe & Londoño-Mesa & Hutchings, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5038.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:1C1E4C7A-2452-47BC-B843-2543135EF780 |
|
persistent identifier |
https://treatment.plazi.org/id/03B887B1-6E29-FFEE-48F7-645F1624940D |
|
treatment provided by |
Plazi |
|
scientific name |
Lanice kellyslateri |
| status |
sp. nov. |
Lanice kellyslateri View in CoL n. sp.
Figures 14–15 View FIGURE 14 View FIGURE 15
Material examined. Holotype. MNHN-IA-TYPE 2026, complete, Northeastern Atlantic , Bay of Biscay , Arcachon Bay, Eyrac pier, 44°39’53”N 01°09’49”W, depth 5 m, October 2016 GoogleMaps . Paratypes. MNHN-IA-TYPE 2027, posteriorly incomplete, Northeastern Atlantic , Bay of Biscay , Arcachon Bay, Eyrac pier, 44°39’53”N 01°09’49”W, depth 5 m, October 2016. AM GoogleMaps W.53328, complete, Northeastern Atlantic , Bay of Biscay , Arcachon Bay , Eyrac pier, 44°39’53”N 01°09’49”W, depth 5 m, October 2016. MNHN-IA-TYPE 2028, posteriorly incomplete, Northeastern Atlantic , Bay of Biscay , Arcachon Bay, Eyrac pier, 44°39’53”N, 01°09’49”W, depth 5 m, October 2016, mounted for SEM GoogleMaps .
Additional material. SMA-Arc-Eyrac-05, posteriorly incomplete, Northeastern Atlantic , Bay of Biscay , Arcachon Bay , Eyrac pier, 44°39’53”N 01°09’49”W, depth 5 m, October 2016, posterior part used for molecular analysis. SMA-Arc-Leg-01, complete, Northeastern Atlantic , Bay of Biscay, Arcachon Bay, Legallais Beach, 44°39’50”N 01°10’29”W, intertidal, December 2017, posterior part used for molecular analysis GoogleMaps .
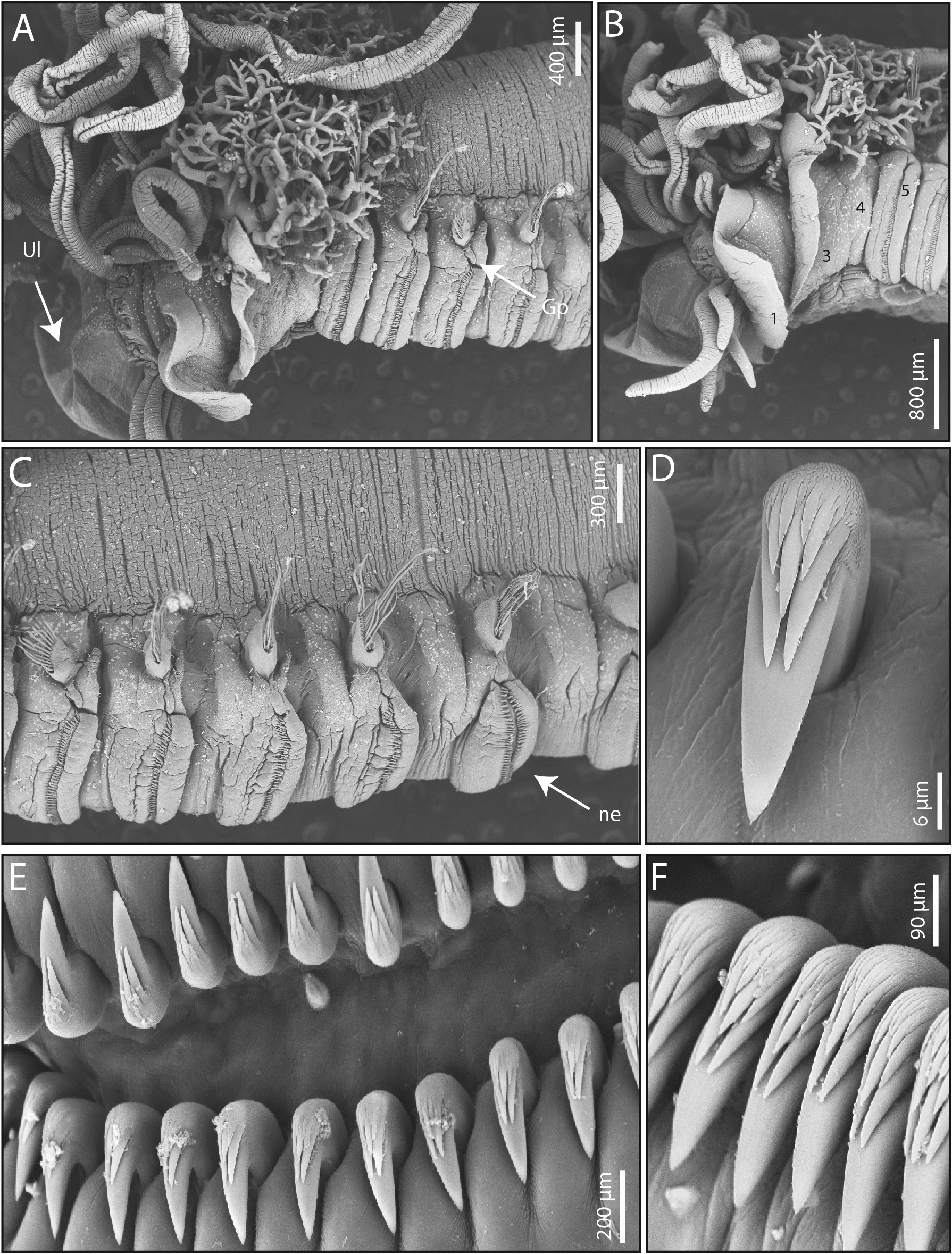
Description. In life, pinkish body, with buccal tentacles translucent; areas around notopodia and neuropodia reddish; lateral lobe on SG I white, on SG III with reddish anterior margin; anterior ventral shields reddish (SG III–XI), following ones blood red (SG XII–XVIII); branchiae blood red ( Fig. 14A View FIGURE 14 ).
Large species, complete holotype 63.3 mm long (61.1 mm), 4.0 mm wide (3.8 mm) for about 130 segments.
Prostomium at base of upper lip, without eyespots, distal part forming shelf-like tentacular membrane from which numerous filiform and deeply grooved buccal tentacles originate ( Figs 14A, C View FIGURE 14 ; 15A–B View FIGURE 15 ). Peristomium well developed, with conspicuous hood-like rectangular upper lip, directed anteriorly, and short and swollen lower lip, partially covered by lobes of SG I ( Figs 14B, D View FIGURE 14 ; 15A–B View FIGURE 15 ).
Segment I with pair of large lobes directed anteriorly and reaching distal part of upper lip; lobes almost oval, pointed anteriorly, with thin undulating margin; continuing across dorsum as narrow crest, connected mid-ventrally by thin convoluted membrane, mid-ventrally indented to partially expose lower lip ( Figs 14A–D View FIGURE 14 ; 15A View FIGURE 15 ). Segment II, short without lateral lobes and laterally covered by lobes of SG III; visible dorsally, with a thick glandular membrane connecting the first pair of branchiae, and ventrally with a pointed anterior projection. Segment III with a pair of large, rectangular lobes, distal margins undulating, with rounded corners, reaching less than mid-length of lobes of SG I laterally; ventral edges fused laterally to first mid-ventral shield. Absence of lateral lobes on SG IV ( Figs 14A–D View FIGURE 14 ; 15A–B View FIGURE 15 ).
Three pairs of arborescent branchiae on SG II–IV, dorsally aligned, with short branchial filaments, with short basal stems ( Figs 14A–C View FIGURE 14 ; 15A–B View FIGURE 15 ); the two first pairs of branchiae connected by a dorsal crest ( Fig. 14B View FIGURE 14 ). Rectangular corrugated mid-ventral shields on SG III–XVIII (XIX), followed by three (two) smaller trapezoidal shields ( Fig. 14D View FIGURE 14 ).
Notopodia beginning on SG IV, extending until SG XX; notopodia of SG IV–XII short, longer from SG XIII, inserted progressively more laterally ( Figs 14B–C View FIGURE 14 ; 15C–D View FIGURE 15 ). Narrowly-winged notochaetae arranged in two rows ( Fig. 14F View FIGURE 14 ), with first row shorter.
Neuropodia present from SG V, as prominent sessile ridges until end of notopodia, reaching mid-ventral shields ( Figs 14A–D View FIGURE 14 ; 15A–C View FIGURE 15 ); from SG XXI as elongate and thin, rectangular pinnules, inserted laterally to mid-ventral groove on posterior segments ( Fig. 15A View FIGURE 15 ). Neurochaetae throughout as short-handled avicular uncini, arranged in back to back double rows on SG XI–XX ( Fig. 15E View FIGURE 15 ). Uncini with triangular heel and rounded prow, with inconspicuous dorsal button, slightly convex base, and main fang surmounted by crest with three rows of secondary teeth, first row with two large teeth, second row with one large and several irregularly-sized teeth, and third row with very small teeth ( Figs 14G–H View FIGURE 14 ; 15D–F View FIGURE 15 ).
Genital papillae on SG VI–XIX, globular, inserted under base of notopodia, slightly displaced dorsally.
Pygidium with eight small globular papillae ( Fig. 14E View FIGURE 14 ).
Tube with sandy-branched projections on anterior margin.
Etymology. This species is named after the “King” Kelly Slater, 11-time Surf World Champion, and also an environmental activist for the protection of the oceans.
Habitat. Sandy beaches, from intertidal to shallow waters (depth 5 m).
Type locality. Eyrac Beach , Arcachon Bay, Bay of Biscay, France ; 44°39’53”N 01°09’49”W.
Distribution. Arcachon Bay ( France) (this study); Scilly Islands ( England) ( Stiller et al. 2020 as L. conchilega , based on molecular results).
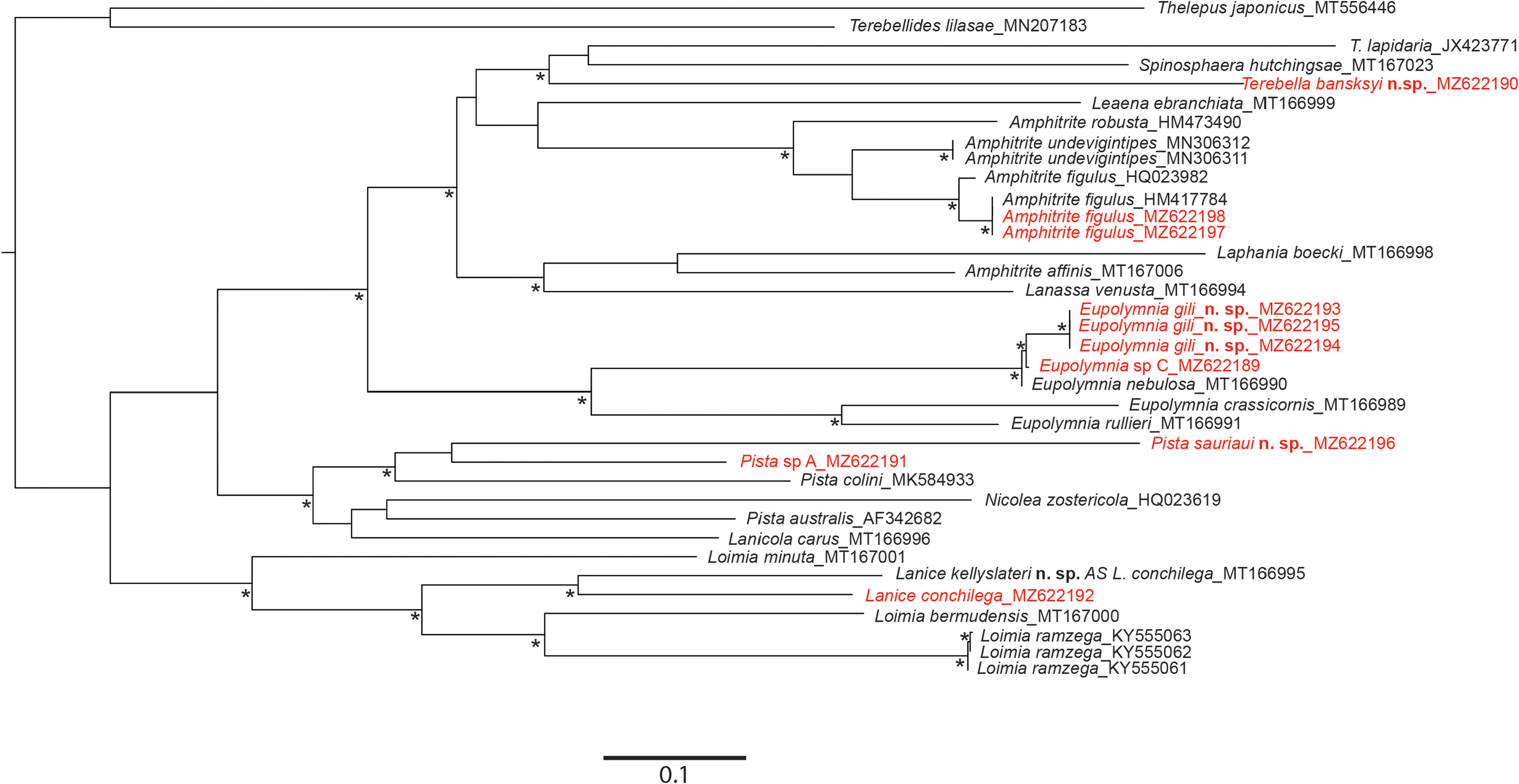
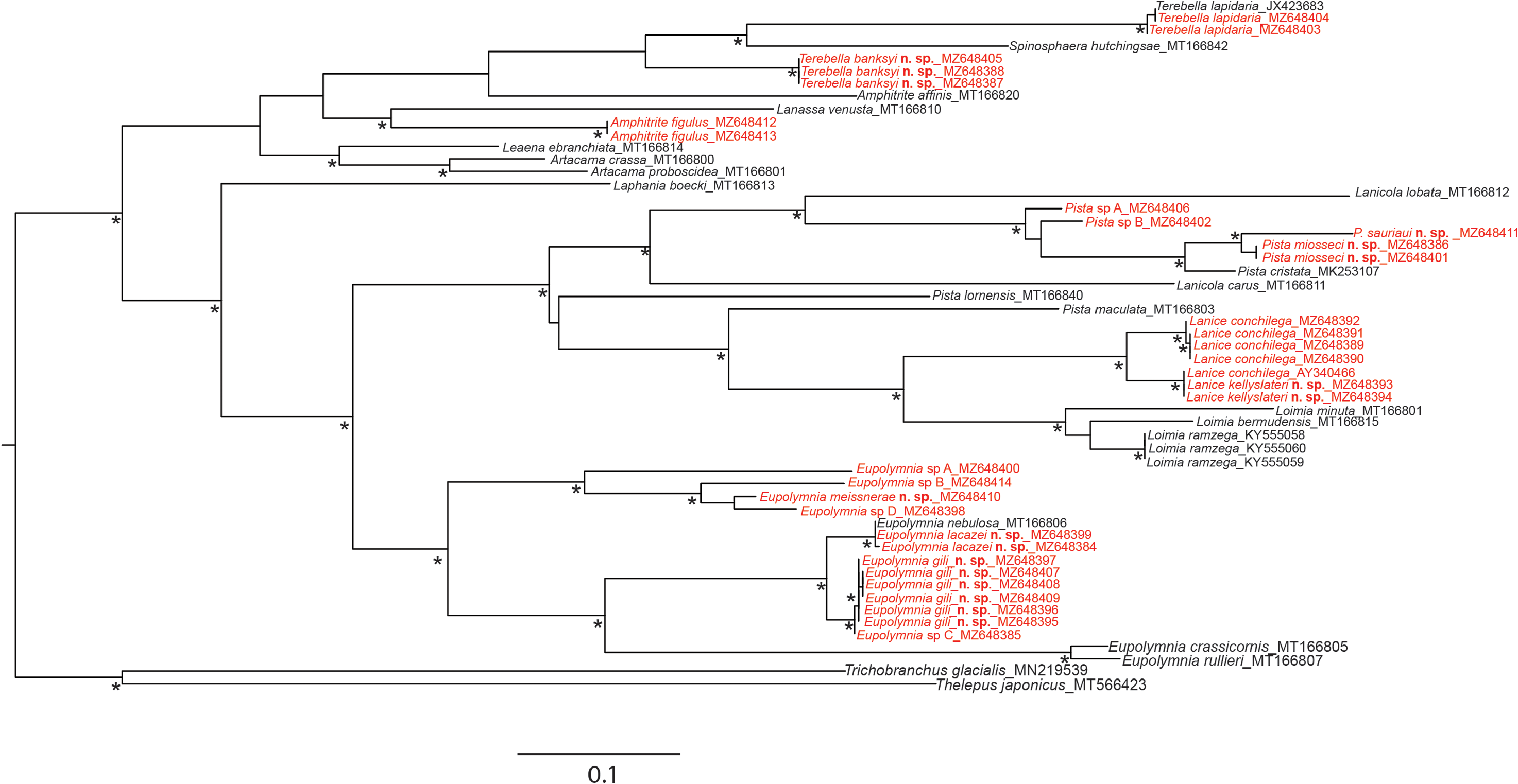
Remarks. Until this study, one single valid species was recorded from European waters: L. conchilega . Molecular analyses ( Figs 26 View FIGURE 26 , 27 View FIGURE 27 ) confirmed the existence of a second cryptic species in France: L. kellyslateri n. sp., which is morphologically very similar. However, L. kellyslateri n. sp. can be differentiated from L. conchilega by the welldefined ventral shields on SG III–IV instead of the presence of ventral shields fused on SG II–IV for L. conchilega , by the upper lip which is never pigmented as in L. conchilega , which is dorsally pigmented with numerous brown spots, by the shape of the neuropodia which are very prominent instead of low sessile ridges as in L. conchilega and by the two first pairs of branchiae connected by dorsal crests, which are absent in L. conchilega .
We have compared L. kellyslateri n. sp. to Terebella pectoralis Quatrefages, 1866 (holotype MNHN-IA-Type 0480), a species described from Atlantic coasts of France and synonymysed with L. conchilega . Even though this material is not in good condition, this species clearly differs from L. kellyslateri n. sp. by the presence of a large ventral homogeneous glandular region instead of well-defined ventral shields present in L. kellyslateri n. sp., by the neuropodia present as low ridges instead of the prominent ones which are present for L. kellyslateri n. sp. and by the presence of short lateral lobes on SG III, with a straight vertical anterior margin instead of the presence of large lobes with undulating margins and rounded corners for L. kellyslateri n. sp. Finally, the tube of T. pectoralis is composed of a smooth membrane and made of gravel and shell fragments while those of L. kellyslateri n. sp. is made of fine sand grains, giving a homogeneous appearance. As pointed out by Quatrefages (1866) and later by Saint-Joseph (1894), the difference of tube morphology within the French Lanice could correspond to different species. The tube of T. pectoralis is indeed very different from L. conchilega and L. kellyslateri n. sp. ones but based on the overall condition of the holotype of T. pectoralis , we are not able to confirm that this species should be considered as valid.
Terebella prudens Quatrefages, 1866 was also described from the same area and later synonymised with L. conchilega . As for T. pectoralis , the description is very brief and lacks important taxonomic details. However, unlike T. pectoralis , no material of T. prudens was deposited in the MNHN collection and thus a comparison was not possible.
Molecularly, L. kellyslateri n. sp. is similar to the L. conchilega identified by Stiller et al. (2020), from Scilly Islands ( England) ( Figs 26 View FIGURE 26 ; 27 View FIGURE 27 ).
Genus Loimia Malmgren, 1866
Type-species: Terebella medusa Savigny, 1822 , by original designation.
Diagnosis. (from Hutchings et al. 2021b). Transverse prostomium attached to dorsal surface of upper lip; basal part as a thick crest, eyespots sometimes present; distal part shelf-like. Buccal tentacles all uniformly cylindrical. Peristomium restricted to lips; lips expanded, relatively short upper lip, hood-like; short, button-like, mid-ventral lower lip. Segment I conspicuous all around, dorsally narrow, with pair of dorso-lateral to lateral lobes extending anteriorly to level of upper lip or beyond, and ventrally, connected to each other by lower membrane across ventrum, partially exposing lower lip. Large lateral lobes present on SG III; SG IV sometimes also with pair of short lateral lobes. Anterior segments highly glandular ventrally, with discrete, smooth to strongly corrugated, rectangular to trapezoidal shields. Three pairs of progressively shorter arborescent branchiae, on SG II–IV, with short main stems. Conical to rectangular notopodia beginning on SG IV, extending for 17 segments, until SG XX; notochaetae all narrowly-winged. Neuropodia beginning on SG V, as low, sessile ridges in conjunction with notopodia and as short pinnules posteriorly; neurochaetae as short-handled uncini throughout, with high, pectinate crest, in partially intercalated to completely separated double rows, arranged in back to back arrangement, from SG XI until end of notopodia. Nephridial papillae on SG III, genital papillae on SG VI–VIII, posterior to notopodia and dorsal. Pygidium smooth to papillate.
Remarks. Recently, Jirkov & Leontovitch (2017) proposed the synonymy of the genera Loimia and Axionice suggesting that the position of the uncini in a back-to-back arrangement, the presence of pectinate uncini, the number of double rows of uncini, and the shape of the lateral lobes were insufficient to consider Loimia as monophyletic. Nevertheless, this synonymization is not followed here, since we consider the genus Loimia as being well defined, with all species having three pairs of arborescent branchiae, uncini in double rows until SG XX, uncini pectinate (at least on the thoracic segments) and lateral lobes well-developed on the SG I. Londoño-Mesa & Glasby (in prep.), after the revision of the existing type material of all currently described species also support Loimia as a valid genus using phylogeny based on morphological information, concluding that the characters listed above as defining the genus are valid and, decline the synonymy suggested by Jirkov & Leontovitch (2017).

| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |