
Lacistema ligiae L. Marinho & K.M. Pimenta, 2021
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.514.2.5 |
|
persistent identifier |
https://treatment.plazi.org/id/03A0B034-B52F-FF89-FF7B-F93DFDA534E8 |
|
treatment provided by |
Felipe |
|
scientific name |
Lacistema ligiae L. Marinho & K.M. Pimenta |
| status |
sp. nov. |
Lacistema ligiae L. Marinho & K.M. Pimenta View in CoL , sp. nov. ( Figures 1 View FIGURE 1 , 2 View FIGURE 2 )
Type:— BRASIL. Bahia: Lençóis, rio Lençóis , 12°32’S 41°23’W, 31 August 1993, fl. fr., L. S GoogleMaps . Funch 678 (holotype: HUEFS barcode HUEFS000032100 About HUEFS !; isotype: ALCB barcode ALCB044377 About ALCB !) .
Diagnosis:— Lacistema ligiae and L. lucidum both have laxly sericeous petioles, leaf blades with simple trichomes on the midvein, and a hairy ovary with a distinct style. Lacistema ligiae can be distinguished by the sinuous secondary veins (vs. straight) with simple, long, and adpressed trichomes abaxially (vs. glabrous), hairy filament (vs. glabrous), and styles not exceeding the height of the stamen (vs. exceeding the height of the stamen).
Description:— Treelets or shrubs, up to 4 m tall, branches with sparsely sericeous indument at the petiole base; terminal stipules glabrescent with simple trichomes at the base and ciliate margin. Leaves alternate, entire; petioles 5–10 mm long, blackish in sicco, with sparsely sericeous indument, trichomes yellowish, simple, long, adpressed. Leaf blade 8–19 × 2–7 cm, chartaceous to subcoriaceous, oblong to elliptic, the apex apiculate, margin serrate for 2/3 distally, revolute, especially at the base, the base attenuate, abaxial surface glabrescent with sparse trichomes, simple, adpressed, smaller than the trichomes of the veins and petioles. Venation: midvein strongly impressed adaxially, prominent, with simple, long, and adpressed trichomes abaxially; secondary veins in 5–6 pairs, strongly impressed adaxially, prominent, with simple, long, and adpressed trichomes abaxially, sinuous, arched near the blade margin, strongly arched near the apex. Spikes axillary, 15–18 per axil, greenish, 4–12 mm long; rachis with trichomes; basal bracts 0.4–0.5 × 0.8–1 mm, naviculate, glabrous, margin erose; bracteoles ca. 3, 0.5–0.6 × 0.1 mm, linear, frequently falcate, glabrous, the apex acute, the margin entire, the base truncate. Flowers monochlamydeous, greenish; sepals 5, 0.4–0.5 × 0.1–0.15 mm, broadly linear to linear, frequently falcate, very brittle, the apex acute, the margin entire, early caducous; stamen 1, 0.7– 0.8 mm long, fused to the nectary, early caducous; filament slightly curved, trichomes present on the distal portion, anthers ca. 0.15 mm long, extrorse, connective thickened, ca. 0.4 mm wide; pollen grains small (polar axis [P]: 17.5–22.5 µm, equatorial axis [E]: 10–15 µm, [DEp]: 12.5–15), with subcircular amb, monads, isopolar, prolate, tricolporate, aperture lalongate, exine microreticulate-perforate, circular puncta, sexine as thick as the nexine; nectary 4-lobed, fleshy; gynoecium 0.5–0.65 mm long, ovary superior, 0.25–0.35 mm long, 3-carpellate, ovule 1 per locule, ovoid, with flattened trichomes on the upper half, styles 0.25–0.3 mm long, glabrous, stigmas 3, not sessile. Capsules 8–10 mm long, ovoid, green to red or purplish when mature, with sparse trichomes when mature. Seed 1, ca. 5 mm long, aril white.
Additional examined specimens (Paratypes):— BRAZIL. Bahia: Ibicoara, fazenda Ponte de Roxo , 12°49’S 41°19’W, 2 December 1986, fl., J GoogleMaps .D.C. Arouck-Ferreira et al. 409 ( CEPEC!, RB). Lençóis, rio Lençóis , 12°33’S 41°26’W, 700 m, 13 September 1993, fl. fr., L. S GoogleMaps . Funch & R . Funch 724 ( HUEFS 2 About HUEFS -sheets!); ibid., na trilha da encosta, 12°33’S 41°26’W, 26 December 1993, fl. fr., L. S GoogleMaps . Funch & R . Funch 743 ( HUEFS!); ibid., 12°31’30”S 41°22’26”W, 500 m, 8 January 1999, fl., R. B GoogleMaps . Santos 9 ( HUEFS!, SPF); 12°33’15”S 41°21’41”W, 550 m, 5 September 2004, fl., A. P. L GoogleMaps . Couto et al. 15 ( HUEFS!); 12°34’S 41°23’W, 600 m, 30 July 2006, fl. fr., A. P. L GoogleMaps . Couto 186 ( HUEFS!, P). Mucugê, Guiné, Vale do Pati , 13°00’S 41°22’W, 8 February 2013, fl. fr., A GoogleMaps . Rapini & P. K . Endress 2003 ( HUEFS!). Palmeiras , km 232 da rodovia BR 242 para Ibotirama, Pai Inácio, 18 December 1981, fl. fr., A. M . Carvalho et al. 978 ( CEPEC!, RB); cachoeirinha Pai Inácio , 12°34’S 41°23’W, 6 April 2004, fl. fr., C GoogleMaps . van den Berg et al. 1403 (HUEFS!).
Etymology:— The specific epithet honors Dr. Lígia Silveira Funch, a professor at the Universidade Estadual de Feira de Santana (UEFS), in Bahia, Brazil. Besides collecting the holotype and various paratypes, she has conducted several studies in Chapada Diamantina vegetation.
Distribution and conservation status:— Lacistema ligiae is restricted to riparian forests ( Fig. 2E View FIGURE 2 ) and semideciduous forest remnants in the northern portion of the Espinhaço Range, in Bahia State ( Fig. 3 View FIGURE 3 ). The conservation status of L. ligiae is here categorized as Endangered (EN) based on an extent of occurrence slightly upper than 1,000 km 2 and an area of occupancy minor than 16 km 2 [B1 + B2a, bii] ( IUCN 2012). Although it falls under a threatened category, all records of L. ligiae have been made within Chapada Diamantina National Park, which is a permanently protected area.
Taxonomic notes:— The first step of the taxonomic key by Sleumer (1980), which separates the species into two distinct groups, is based on the sections proposed by Chirtoiu (1918): Lacistema sect. Lacistema (“ Eulacistema ” in Chirtoiu 1918: 329), with a very short or practically no style and sessile or nearly sessile stigmas; and Lacistema sect. Stylolacistema Chirtoiu (1918: 329) , with a distinct style at least 0.3 mm long ( Sleumer 1980). Among the seven Lacistema species with distinct styles, including L. ligiae , L. polystachium Schnizlein (1857: 284) is endemic to the Amazonian forest, L. grandifolium Schnizlein (1857: 284) and L. pubescens Martius (1824: 155) have leaves that are densely pubescent to tomentulose all over the undersurface, and L. serrulatum Martius (1824: 156) has leaves distinctly serrate on the upper 2/3 of the lamina. Thus, L. ligiae is compared to L. lucidum and L. robustum .
In the Flora da Bahia recently published by Marinho & Amorim (2015), specimens of Lacistema ligiae were labeled as L. lucidum and the first records of L. lucidum for the Northeast Region of Brazil. These species are similar but occur far apart: L. ligiae is endemic to Bahia State, in areas of riparian ( Fig. 2E View FIGURE 2 ) and semideciduous forest; and L. lucidum occurs in the subtropics in the states of Paraná and São Paulo ( Fig. 3 View FIGURE 3 , Sleumer 1980, Torres & Ramos 2005, Marquete & Medeiros 2021), in areas of restinga and Atlantic Forest ( Torres & Ramos 2005). Morphologically, L. ligiae can be distinguished by the leaf blades that reach 19 cm long and 7 cm wide (vs. 14 cm long and 4 cm wide in L. lucidum ) ( Fig. 2B View FIGURE 2 ) and leaf blades with sparsely distributed trichomes, which are longer and denser on the veins (vs. glabrous with sparse trichomes only on the midrib in L. lucidum ) ( Fig. 4A View FIGURE 4 ). Additionally, in L. lucidum the styles exceed the height of the stamen ( Torres & Ramos 2005), which does not occur in L. ligiae .
Some specimens of Lacistema ligiae were identified as L. robustum , a species widely distributed in humid forests from the state of Pernambuco to the state of Espírito Santo ( Fig. 3 View FIGURE 3 ). These species can be especially differentiated by vegetative characters: petiole with sparse trichomes, margin of leaf blade serrate at the distal portion and revolute on the proximal portion in L. ligiae (vs. glabrous petiole, margin of leaf blade entire, not revolute in L. robustum ) (see Fig. 4 View FIGURE 4 ). They also occur far apart from each other in the state of Bahia; L. robustum grows in areas closer to the coast ( Fig. 3 View FIGURE 3 ).
Micromorphological discussion:— The leaf vein patterns of the Lacistema species analyzed have secondary veins in low numbers (ca. 7 pairs) that form uniform and acute angles with the midvein. The secondary spacing decreases proximally and there are no intersecondary veins. The tertiary veins are percurrent sinuous, crossing between adjacent secondaries and changing the direction of the curvature. The leaf clarification analyses showed that the veins of L. ligiae are eucamptodromous, and the secondary veins are extremely ascending and sinuous ( Fig. 4A–B View FIGURE 4 ). In L. lucidum , the leaf vein pattern is eucamptodromous and becomes brochidodromous distally ( Fig. 4C View FIGURE 4 ). In this case, the proximal secondaries are eucamptodromous, but distal secondaries form loops of secondary veins ( Ellis et al. 2009). These loops are present in the leaves of L. robustum , which is simple brochidodromous ( Fig. 4E View FIGURE 4 ). The secondary veins of L. lucidum and L. robustum are straight like those of L. ligiae , which is a striking feature in the last species.
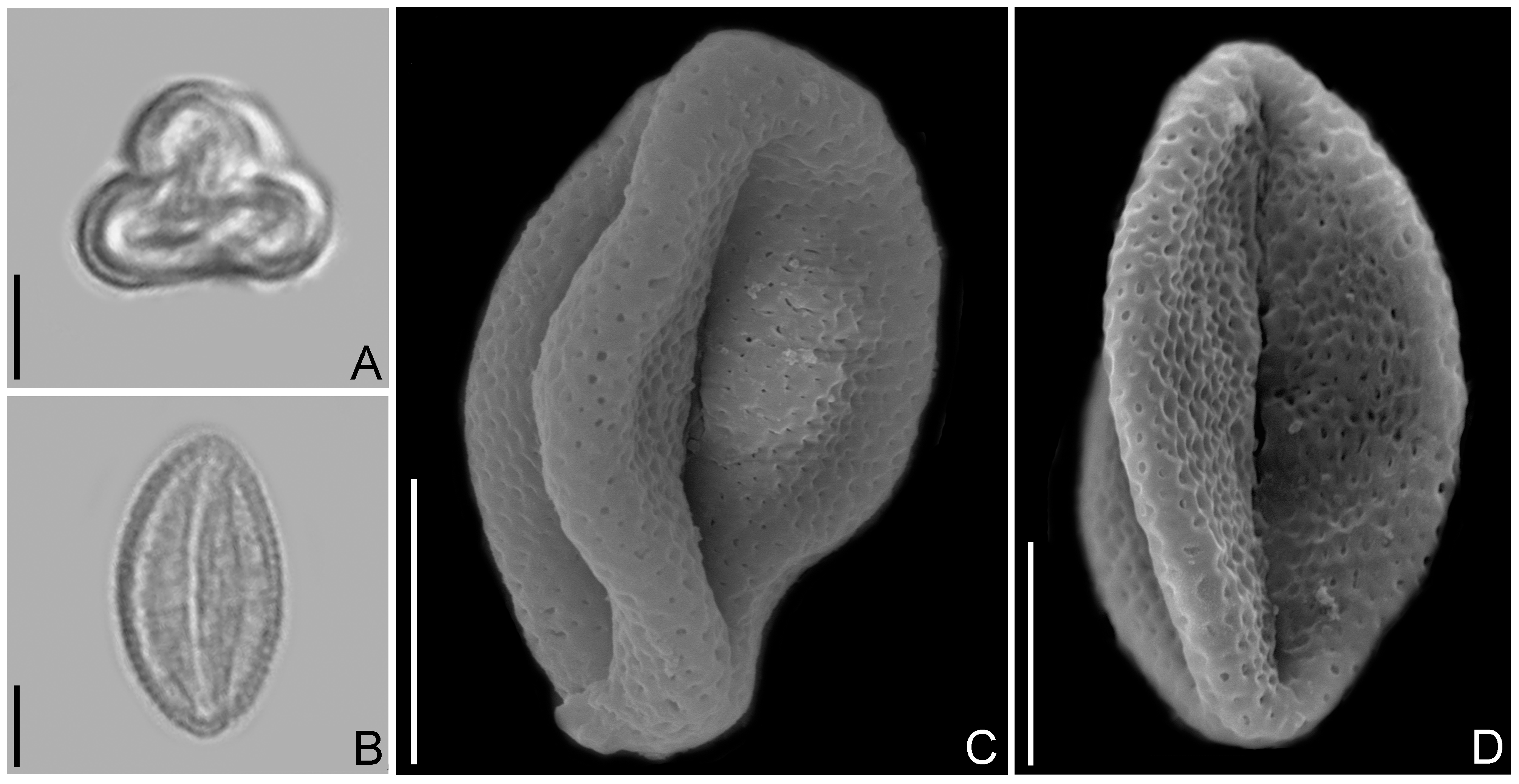
Few studies include pollen grain descriptions of Lacistemataceae species. Here, we present the first scanning electron micrographs of Lacistema pollen ( Fig. 5 View FIGURE 5 ). Lorente et al. (2017) described the pollen grains of L. recurvum Schnizlein (1857: 283) (a synonym of L. aggregatum (P.J. Bergius [1772: 131] Rusby [1907: 447], the type of the genus) as small, monads, tricolporate, trilobate to circular, prolate to subprolate, microreticulate and homobrocate. The pollen grains of L. aggregatum were also described by Leal et al. (2011) as monads, tricolporate and trilobate to circular, but pointed out the grains as psilate. Since the species has a wide distribution and an extensive list of synonyms (see Sleumer 1980), it would be important to investigate further specimens. Ybert et al. (2017) found pollen grains monads, tricolporate with subtriangular amb and microreticulated exine for L. pubescens . Pollen grains from both, L. aggregatum and L. pubescens , have similarities with the pollen of L. ligiae and L. lucidum . In the last two, the reticulum decreases towards the margo and apocolpium ( Fig. 5 View FIGURE 5 ). Although small, monads and tricolporate, it is still possible to distinguish pollen grains of L. ligiae ( Fig. 5A–C View FIGURE 5 ) from L. lucidum ( Fig. 5D View FIGURE 5 ), which are microreticulate (vs. microreticulate-perforate in L. ligiae ).




| L |
Nationaal Herbarium Nederland, Leiden University branch |
| S |
Department of Botany, Swedish Museum of Natural History |
| HUEFS |
Universidade Estadual de Feira de Santana |
| ALCB |
Universidade Federal da Bahia, Campus Universitário de Ondina |
| J |
University of the Witwatersrand |
| CEPEC |
CEPEC, CEPLAC |
| RB |
Jardim Botânico do Rio de Janeiro |
| R |
Departamento de Geologia, Universidad de Chile |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| SPF |
Universidade de São Paulo |
| A |
Harvard University - Arnold Arboretum |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| K |
Royal Botanic Gardens |
| M |
Botanische Staatssammlung München |
| C |
University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |