
Laberia palliata Stål, 1866
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.836.1913 |
|
publication LSID |
lsid:zoobank.org:pub:EAD4EC62-D847-4A91-86F1-511EAA5FC6B8 |
|
DOI |
https://doi.org/10.5281/zenodo.7055014 |
|
persistent identifier |
https://treatment.plazi.org/id/03D9E940-FFAE-FFD6-FDB6-F722CE5EFA9B |
|
treatment provided by |
Felipe |
|
scientific name |
Laberia palliata Stål, 1866 |
| status |
|
Laberia palliata Stål, 1866 View in CoL
Figs 3–16 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig
Perinetia reticulata Lallemand & Synave, 1954: 82 ; syn. nov.
Aylaella reticulata View in CoL – Demir & Özdikmen 2009: 271.
Type material
Holotype of Laberia palliata Stål, 1866 ( Fig. 3 View Fig ) LOCALITY UNKNOWN • ♀; “ Mauri- / tius .”, “ Stevens.”, “Typus”, “140 / 64”, “ Laberia pallida Stål / Holotype ( Flatidae )”, “ NHRS-GULI / 000006158 ”; NHRS .
Holotype of Perinetia reticulata Lallemand & Synave, 1954 ( Fig. 4 View Fig ) MADAGASCAR • ♂; “MADAGASCAR / Rég. Périnet / G. Olsoutiett col. 193”, “TYPE”, “ V. Lallemand et / H. Synave det., 195? / Perinetia g. n. / reticulata sp. n. ”, “Museum Paris / MNHN ( EH) / 19341”; MNHN .
Additional material examined
MADAGASCAR – Alaotra-Mangoro Region, Moramanga District • 1 ♂; “ Analamazotra / Perinet, Madag / OЛСуФЪЕва XI.930”; ZIN • 1 ♀; “Périnet / Madagascar / Seyrig 12.II. 931”; ZIN • 1 ♀; “ Madagascar / province de Toamasina / Andasibe , 1049 m / S18°53,410, E48°22,881 ”, “Museum Paris / frt humid, brd piste nicke / 04-XI-2011 / Th. Bourgoin Rèc. ”, “Museum Paris / MNHN (EH) / 19347”; MNHN GoogleMaps • 1 ♂, 1 ♀; “ Madagascar –CE / border of Andasibe N.P. / “Parc de Orchidés”/ at light; 3.-6. xi.2010 / P. Bănař & local coll. lgt.”, “Collectio / Moravské museum / Brno”; MMBC. – Analamanga Region, Manjakandriana District • 1 ♀; “ MADAGASCAR CENTRE / EST LAC MANTASOA / AMBOHIBOATAVO ”, “ III-1973 / A. Peyrieras ”, “Museum Paris / MNHN ( EH) / 16576”; MNHN • 1 ♀; “ MADAGASCAR CENTRE / EST LAC MANTASOA / AMBOHIBOATAVO ”, “ III-1973 / A. Peyrieras ”, “Museum Paris / MNHN ( EH) / 16577”; MNHN. – Vatovavy Fitovinany Region, Ifanadiana District • 1 ♂; “ Madagascar –CE / Ranomafana N.P. ; 11.– / 18.xi. 2010, 958m / S21°15′22.6″ E47°25′17.8″ / at light; P. Bănař lgt.”, “Collectio / Moravské museum / Brno”; MMBC. – Vatovavy Fitovinany Region: Ifanadiana District, Ranomafana commune GoogleMaps • 1 ♀; “COLL. MUS. CONGO / Madagascar: Nandihizina / XII – 1937, J. Vadon ”, “ Laberia paliata Stål , det / RGFennah”; MRAC .
Redescription
MEASUREMENT. Total length 15–18 mm.
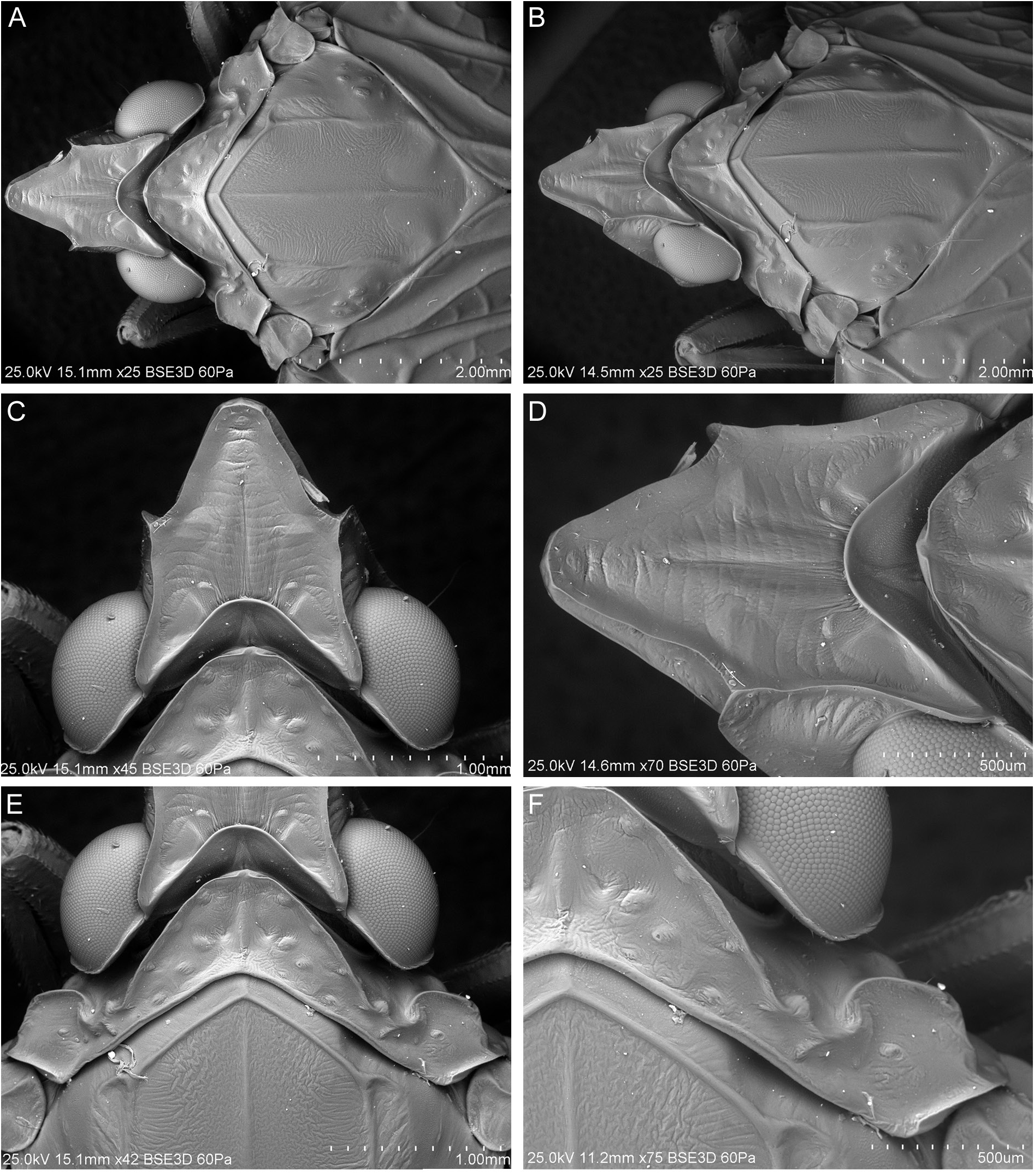
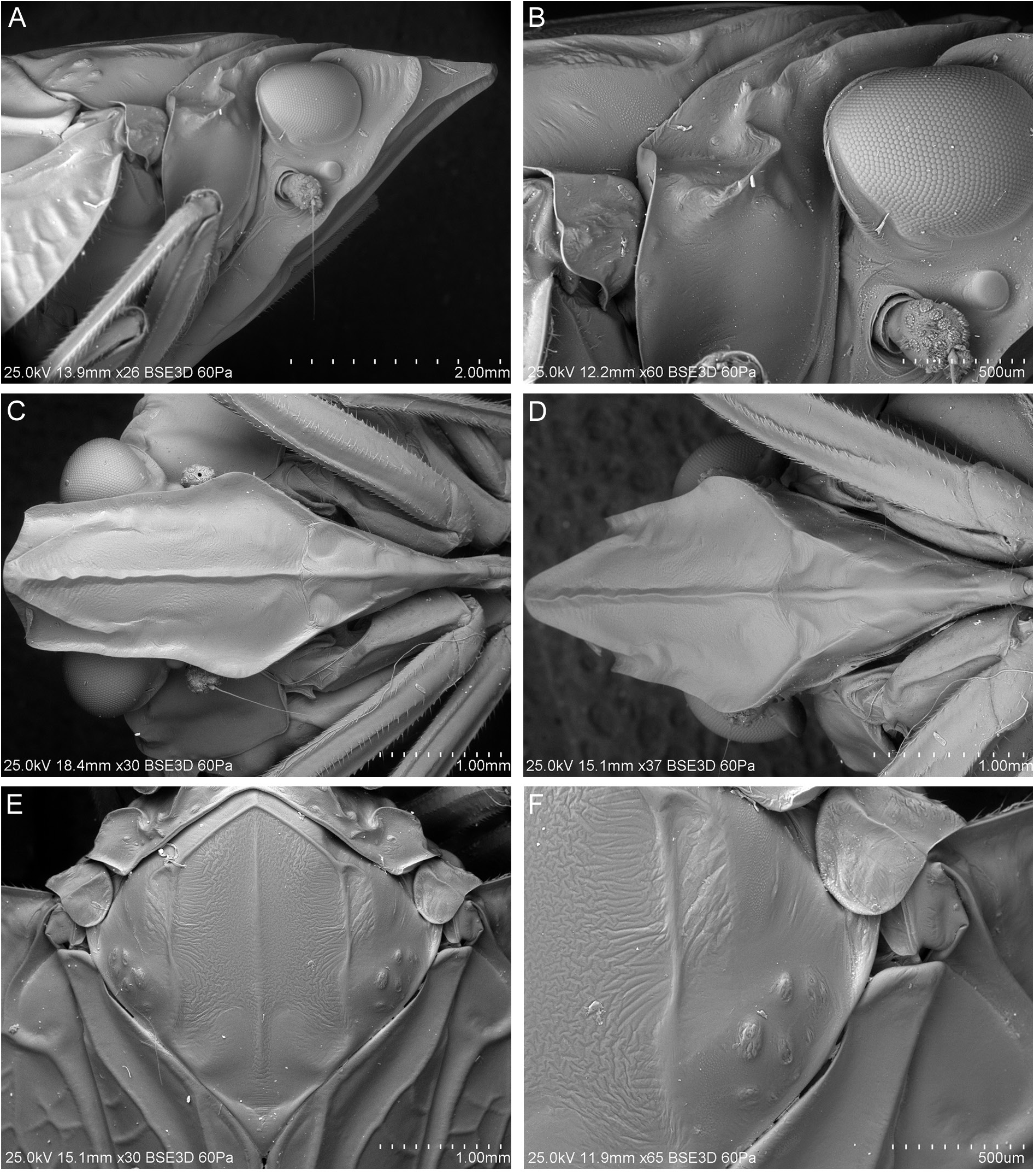
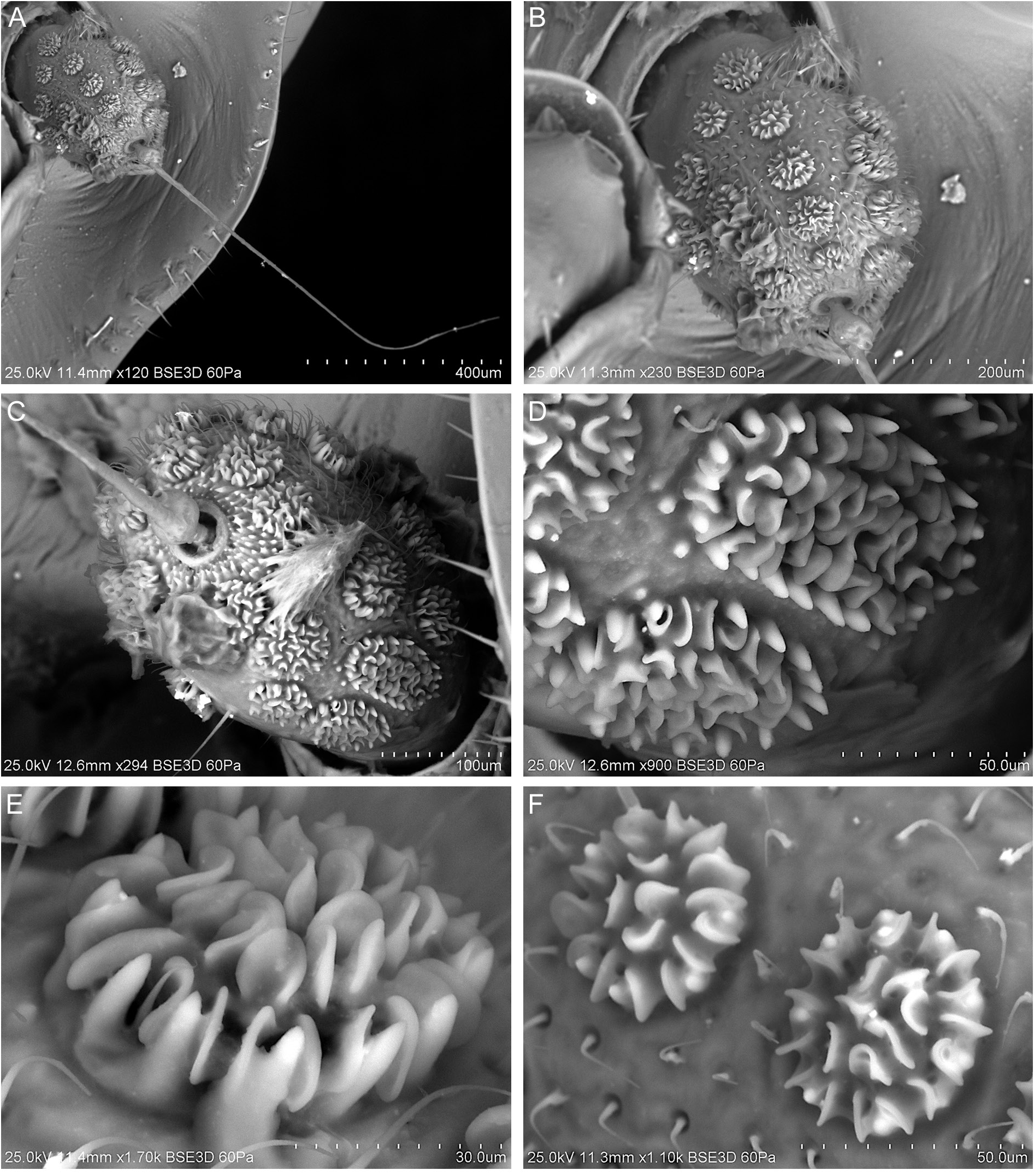
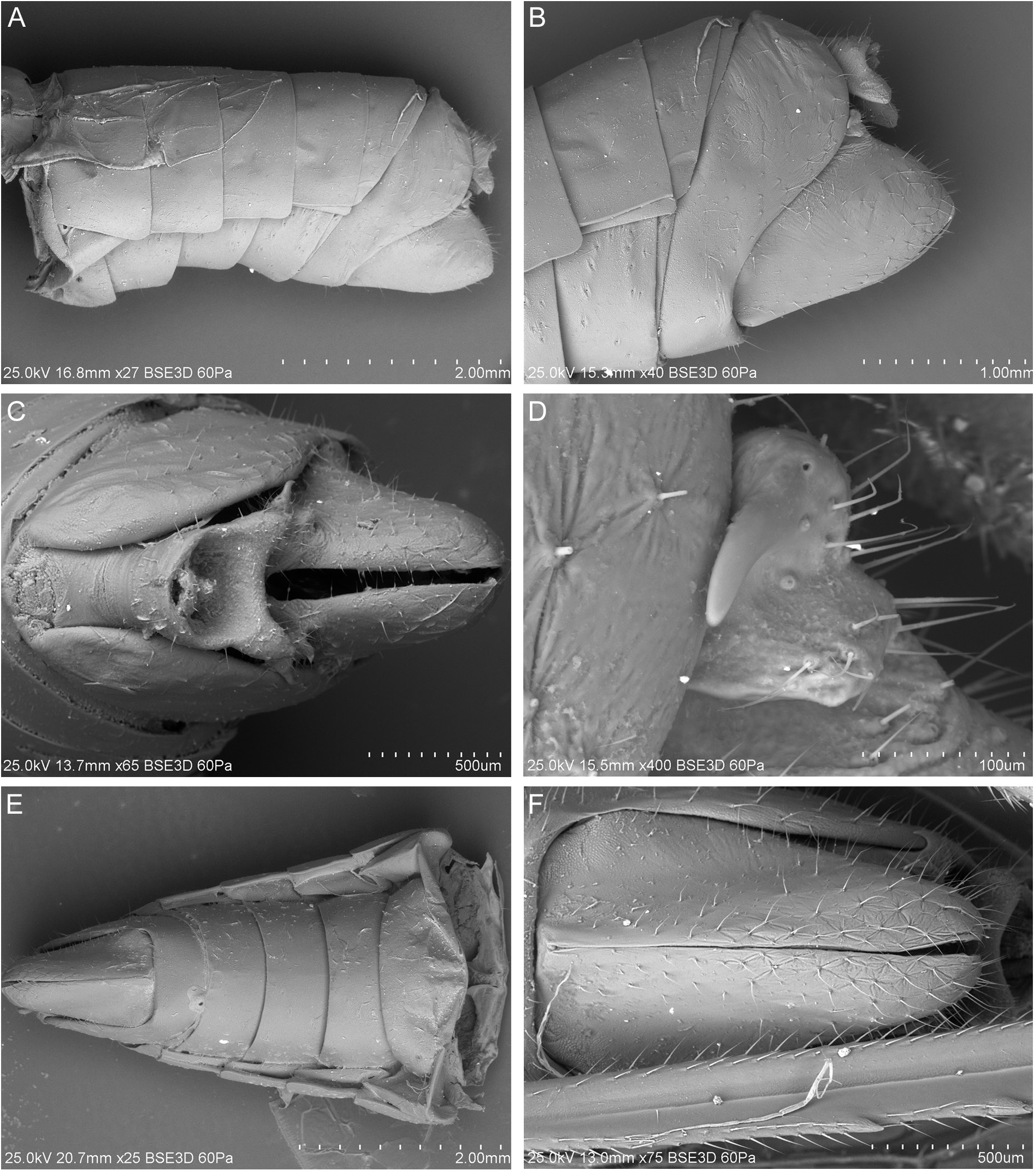
HEAD. Head with compound eyes narrower than pronotum ( Fig. 6A–B View Fig ). Disc of vertex, pronotum and mesonotum in one plane ( Fig. 7A View Fig ). Vertex ( Fig. 6A–D View Fig ): ratio A/B = 1.0–1.1, with margins slightly elevated, carinate; anterior margin of vertex triangularly produced, apex of vertex obtusely angled; posterolateral margins of vertex slightly diverging posteriad; posterior margin distinctly triangularly incised, to the level of anterior margins of compound eyes; disc of vertex flat, without median carina, with incomplete furrow spreading from posterior margin to anterior ¼ of vertex length in mid line. Frons ( Fig. 7C–D View Fig ): ratio C/E = 0.41–0.49; D/E = 1.58–1.67; frons longer than wide, widest below level of antennal bases; anterior margin prolonged, lateral margins carinately elevate; disc of frons with three carinae, median carina reaching frontoclypeal suture, lateral carinae not exceeding level of lower margins of compound eyes, median and lateral carinae connected at base. Postclypeus with median and lateral carinae; anteclypeus medially carinate; anterior margin of preocular field covered with row of short setae ( Fig. 7C–D View Fig ). Rostrum reaching between metacoxae; apical segment shorter than subapical one. Compound eyes ellipsoidal, posteriorly with narrow callus. Ocelli distinct. Antennal fovea emarginate, posteriorly elevated, shifted to posterior margin of gena. Scape short, cylindrical. Pedicel bulbous, slightly longer than wide, arista about 3 times as long as pedicel; antennal plate organs evenly distributed on pedicel, organized in rows, reaching base of pedicel. Plate organs of crenelated plate type in the merged-tip type group, with ear-like lobes paired organized on the external circle of them, isolated in the internal area ( Fig. 8 View Fig ).
THORAX. Pronotum ( Figs 6 View Fig , 7E–F View Fig ): ratio F/B = 0.44–0.54; disc of pronotum arcuate, with median carina and lateral impressions, lateral carinae not reaching posterior margin, anterior angle incised between compound eyes, exceeding anteriorly half of compound eyes length; posterior margin of pronotum widely triangularly incised, incision reaching level of posterior margins of compound eyes; lateral lobes with curved anteriad; distinctly elevated postocular carinae, not reaching posterior margin of pronotum; lateral carina and horizontal carina of side of pronotum absent. Mesonotum ( Figs 6A–B, E–F View Fig , 7E–F View Fig ): ratio G/F = 5.3–6.13, G/B+F = 1.80–2.08, G/H = 0.99–1.03; in mid line about as long as wide, diamond-shaped, with disc flat and lateral portions declivous, with five parallel carinae; median carina reaching almost to scutellum, lateral carinae reaching posterior margins, anterolateral carinae pronounced at base than obsolescent posteriad; mesoscutellar groove arcuate posteriad. Tegula with two parallel carinae ( Fig. 9A View Fig ).
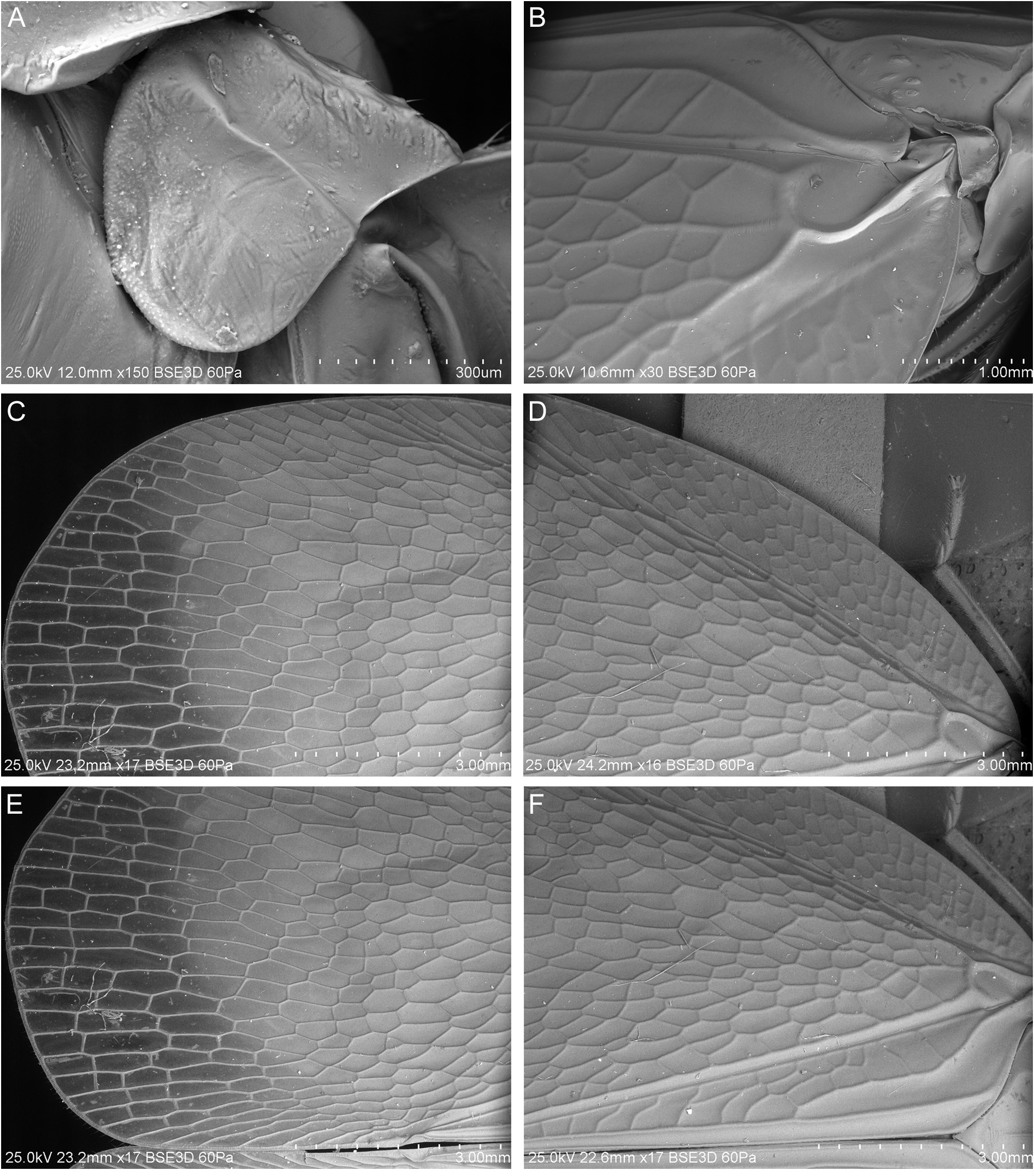
TEGMINA ( Figs 9B–F View Fig , 10A–D View Fig ). Ratio I/J = 1.86–2.02; membranous, with dense network of veins and veinlets. Costal margin curved at base, arcuate to anteroapical angle; anteroapical angle widely arcuate, posterior margin arcuate, posteroapical angle widely obtuse; tornus straight; apex of clavus reaching ⅔ of tegmen length. Costal area present, wide, with apex reaching almost to ⅔ of tegmen length, merely basad of apex of clavus; costal area with prominent, reticulate network of veinlets. Basal cell rounded, slightly longer than wide. Stems ScP+R, MP and CuA leaving basal cells separately, in ScP+R→MP→CuA forking sequence on corium. Stem ScP+R with short stalk, shorter than basal cell, both branches ScP+RA and RP with few furcations, together with veinlets resulting in reticulate pattern, ultimate terminal of RP reaching anteroapical angle. Common stalk of MP about as long as basal cell, both branches MP 1+2 and MP
3+4 with number of furcations, together with veinlets resulting in reticulate pattern, the earliest terminal of MP 1+2 reaching anteroapical angle, the ultimate terminal of MP 3+4 exceeding posteroapical angle, reaching tornus. Common stem CuA distinctly longer than basal cell; branch CuA 1 weaker, with a few furcations, reaching tornus with three terminals, branch CuA 2 stronger, parallel to claval suture and vein CuP, reaching margin with single terminal. Claval suture and vein CuP distinct. Claval veins Pcu and A 1 fused in basal half of clavus length, joined vein Pcu+A 1 reaching apex of clavus; irregular veinlets between CuP and Pcu present. Costal cell narrow, with a few veinlets between stems Pc+CP and ScP+RA. Nodal line not materialized. Chitinized pterostigmal area absent. Veinlet icu present, connecting CuA 2 and apex of clavus. Veins with short, scarce setae on ventral surface.
HIND WING. Membranous, costal margin slightly curved at base, then almost straight, slightly concave at level of wing coupling apparatus, anteroapical angle widely rounded, apical margin distinctly curved, posteroapical angle widely rounded, anal lobe widely rounded. Basal cell slightly longer than wide, stems ScP+R and M leaving basal cell with a short common stalk. Stem ScP+R forked at level of wing coupling apparatus, ScP+RA 1 reaching margin distinctly basad of anteroapical angle, with 3 terminals; single rarp veinlet present. RP forked apicad of transverse veinlet rp-mp, about at the level of ending RA vein; RP with 4–6 terminals reaching margin basad of and at anteroapical angle. Transverse veinlets rp-mp 3–4. Stem MP forked usually about level of first fork of RP, with 4–5 terminals, single imp vein present between MP 1+2 and MP 3+4. Stem CuA forked distinctly basad of stem ScP+R forking, before half of wing, multifurcation with 11–16 terminals and with 10–12 icu veins. Stem CuP single, stem Pcu subparallel to CuP in basal portion, apically curved towards CuP. Veins with short scarce setae on ventral surface.
LEGS ( Figs 10E–F View Fig , 11 View Fig ). Profemur slightly flattened, with rows of short setae along margins, subquadrate in cross section, with rows of short setae along margin; basiprotarsomere as long as midprotarsomere, apical protarsomere slightly longer than cumulative length of basi-and midprotarsomeres; tarsal claws distinct, arolium bilobate, wide. Mesofemur ( Fig. 10E View Fig ) slightly longer than profemur, subquadrate in cross section, with rows of short setae along margins, basimesotarsomere ( Figs 10E View Fig , 11A View Fig ) as long as midmesotarsomere, apical mesotarsomere slightly longer than cumulative length of basi-and midmesotarsomeres; tarsal claws distinct, arolium bilobate, wide. Metafemur ( Fig. 11B View Fig ) shorter than metatibia, metatibia with 5–7 lateral spines, subquadrate in cross sections, with margins covered with rows of short setae, with asymmetrical 8 apical teeth ( Fig. 11B–D View Fig ). Basimetatarsomere ( Fig. 11C–D View Fig ) long, about as long as cumulative length of mid- and apical metatarsomeres, apical margin incised, with 12 apical teeth, with subapical setae, except the external ones; midmetatarsomere long, lateral teeth distinct, ventromedian margin arcuately convex, with bunch of setae; apical metatarsomere with rows of longitudinal setae; tarsal claws distinct, arolium bilobate.
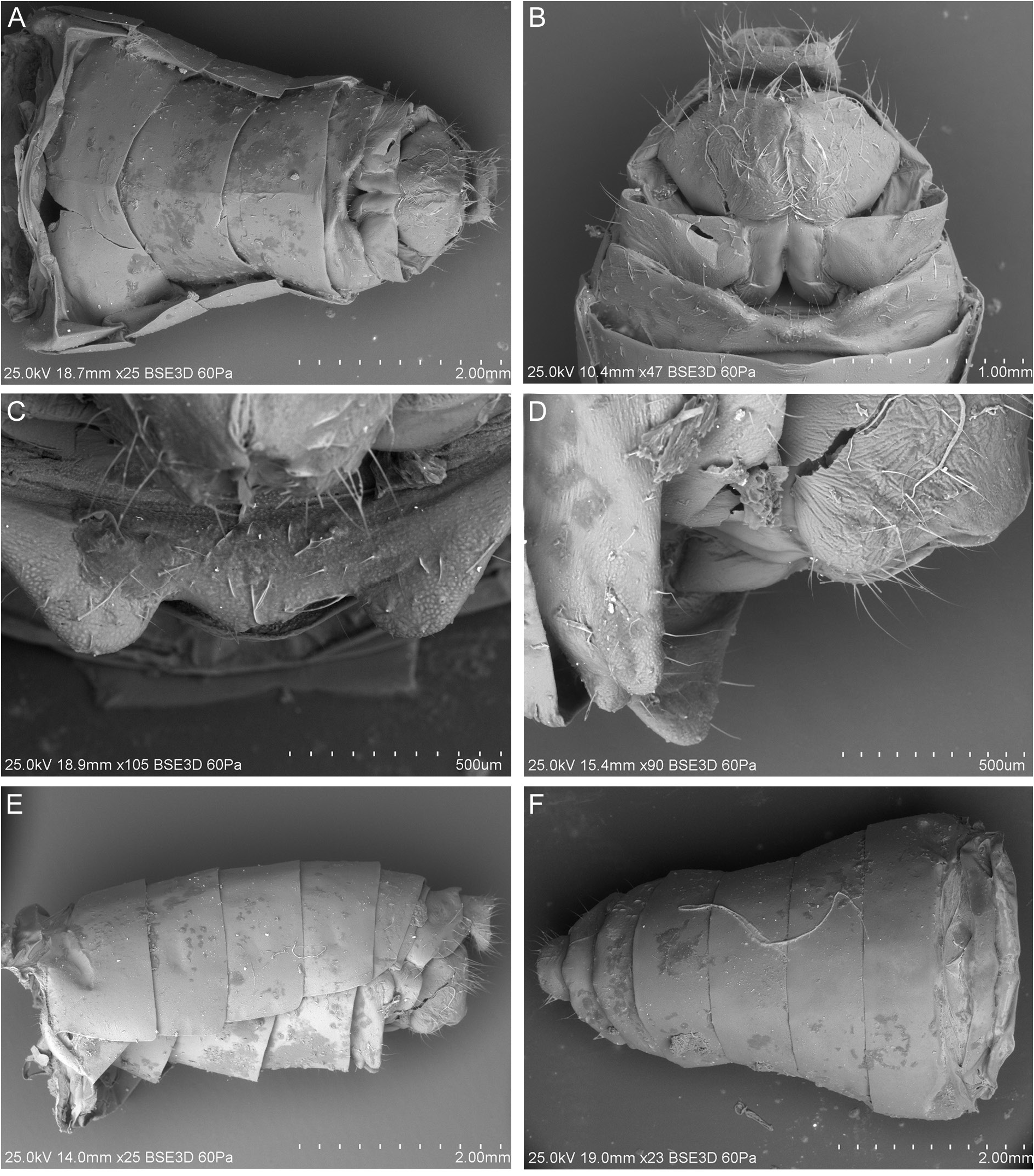
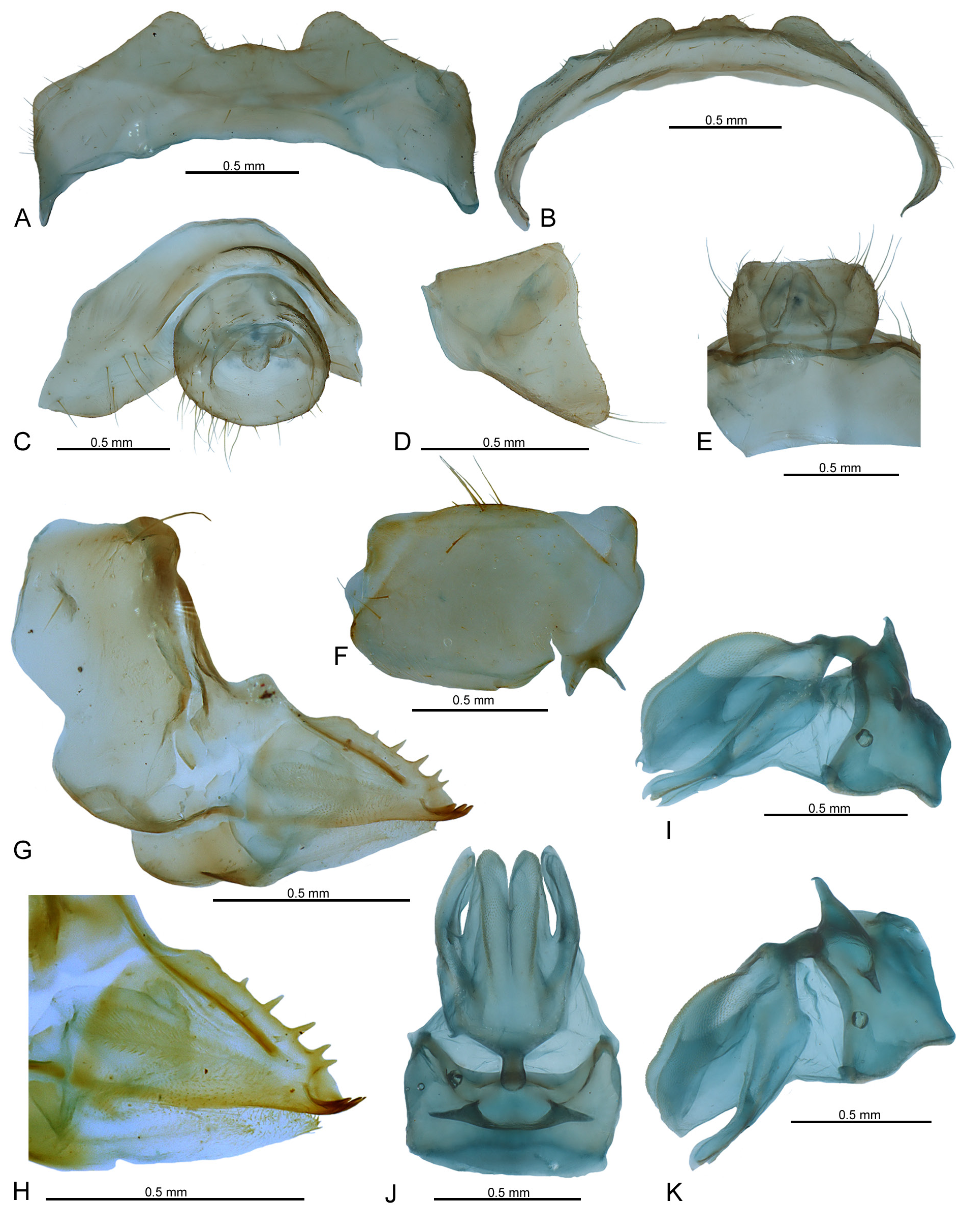
MALE TERMINALIA ( Figs 12–13 View Fig View Fig ). Anal tube ( Figs 12A–C View Fig , 13A View Fig ) tubular, slightly longer than wide with posterior margin concave in dorsal view, produced ventrally in lateral view. Pygofer ( Figs 12A–B View Fig , 13C View Fig ) with upper part distinctly wider than ventral, posterior margin strongly sinuated, without any processes. Corpus connectivi ( Fig. 13D View Fig ) short and robust; alae connectivi large, bearing well developed crista. Periandrium ( Fig. 13D–F View Fig ) covering half of aedeagus, slightly asymmetrical, with two pairs of broad spine-like processes; dorsal processes bigger than ventral ones; ventral basal spine-like process on right side and a short membranous bulb, covered with minute teeth. Aedeagus ( Fig. 13G–H View Fig ) distinctly longer than periandrium, long and narrow, apically, with a pair of dorsal sclerotized strong spines, surrounding a pair of hook-like longer processes.
FEMALE TERMINALIA. Pregenital sternite ( Figs 14A–D View Fig , 16A–B View Fig ) well developed with posterior margin with huge submedian protruding lobes with margin between them weakly arcuate. Anal tube ( Figs 14A View Fig , 15A–D View Fig , 16C–E View Fig ) round, short: shorter than wide in dorsal view. Paraproct small, apical margin widely rounded, reaching posterior margin of anal tube. Epiproct slightly shorter than paraproctal lobe. Gonoplac ( Figs 14A–B View Fig , 15E–F View Fig , 16F View Fig ) quadrangular, slightly sclerotized. Gonospiculum bridge short and robust ( Fig. 16I–K View Fig ). Gonocoxa VIII ( Fig. 16G–H View Fig ) trapezoidal, with a deep concave incision on its anterior margin. Endogocoxal lobe ( Fig. 16G–H View Fig ) bearing a membranous endogonocoxal process with minute apical teeth, slightly shorter than the anterior connective lamina. Anterior connective lamina with some strong 5 subterminal and 3 apical well sclerotized teeth ( Fig. 16G–H View Fig ). Fibula anterior slightly curved. Gonapophysis IX elongated, membranous with median lobes united medially, straight, densely covered dorsally with minute, scale-like denticulations, almost as long as lateral lobes, which bear the posterior fibulae; lateral margin with a long, developed finger-like lobe, covered dorsally with minute, scale-like denticulations.
COLORATION ( Figs 3A–C View Fig , 4A–F View Fig , 5 View Fig ). General coloration green. In dry and older specimens, general coloration reddish-brown or brown. Hind wing membranous, transparent, with one dark, round spot on anal area. Abdomen in ʻfreshʼ specimens with orange tergites and sternites yellowish, yellowish-brown to brown in oldest specimens.
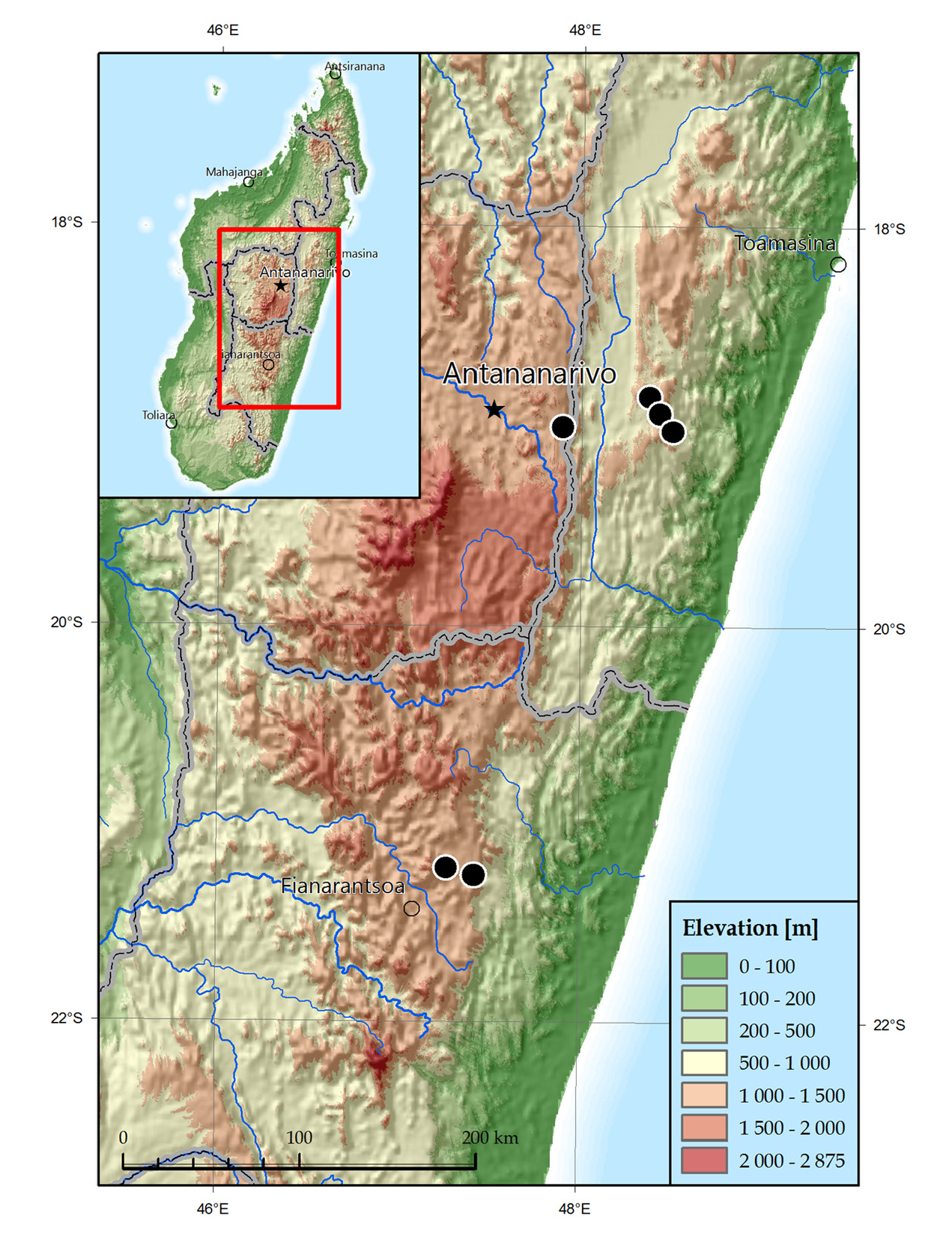
Distribution ( Fig 17A View Fig )
Madagascar (East): Alaotra-Mangoro Region: Moramanga District: Andasibe commune; Analamanga Region: Manjakandriana District, Mantasoa commune; Haute Matsiatra Region: Ambohimahasoa District, Morafeno commune; Vatovavy Fitovinany Region: Ifanadiana District, Ranomafana commune.
Note
The genus Laberia was described by Stål (1866), based on single female specimen labelled as originating from Mauritius ( Fig. 3D View Fig ), as a new genus belonging to Ricaniidae . This seems to be a case of mislabelling, as since the original description it was never reported on Mauritius, and all specimens available came from Madagascar. Melichar (1898: 296) moved this genus to Nogodinidae , and Fennah (1978: 118) placed it in the tribe Bladinini Kirkaldy, 1907 , subtribe Gaetulina Fennah, 1978 . The subsequent changes in placement of Laberia in Tropiduchidae resulted from taxonomic views and opinions concerning Gaetuliini (see above). However, the type material of Laberia has never been re-examined since its description and its characters and taxonomic status needed to be reconciled with the taxonomic changes that took place around the families Nogodinidae-Tropiduchidae and their respective delimitation. A few additional specimens of the species were found dispersed in various collections enabling this revisionary study that led to our separation of the genus into a new tropiduchid Elicinae tribe, Laberiini trib. nov. Additionally, during this study, we also discovered that another ‘mythical’ taxon from Madagascar, Perinetia reticulata Lallemand & Synave, 1954 , originally described as a species of a monotypic genus of the family Acanaloniidae Amyot & Serville, 1834 , was conspecific with Laberia palliata Stål, 1866 .
The generic name Perinetia was proposed by various authors several times for various species of animals. Currently, all these usages have been replaced by new names. Hence, the generic name Perinetia becomes again available for the purposes of taxonomy. However, the oldest usage of this name for a genus is now believed to be a junior objective synonym. All subsequent proposals with Perinetia as generic name have already been replaced.
The usages of the generic name Perinetia are presented below:
1936 Perinetia Collenette View in CoL : 165 [ Insecta: Lepidoptera : Lymantriidae ]; type species: Perinetia leucocloea Collenette, 1936: 166 , pl. 12, fig. 8; by original designation.
Remark. Perinetia Collenette, 1936 has priority, while the other names should be treated as junior homonyms (Article 60 of the International Code of Zoological Nomenclature – ICZN 1999). Perinetia Collenette, 1936 is a junior subjective synonym of Stenaroa Hampson, 1910: 444 [ Insecta: Lepidoptera : Lymantriidae ].
1952 Perinetia Seyrig View in CoL : 193. [ Insecta: Hymenoptera : Ichneumonidae ]; type species: Perinetia nigrifacies Seyrig, 1952: 194 View in CoL ; by original designation.
Remark. This name was proposed to be replaced by Madagascesa Koçak & Kemal, 2008: 6 .
1954 Perinetia Lallemand & Synave View in CoL : 81 [ Insecta: Hemiptera : Acanaloniidae ]; type species: Perinetia reticulata Lallemand & Synave, 1954: 82 .
Remark. This name was proposed to be replaced by Aylaella Demir & Özdikmen, 2009: 271 .
1959 Perinetia Barnard View in CoL : 81 [Crustacea: Malacostraca: Philosciidae ]; type species: Philoscia (Perinetia) reducta Barnard, 1958: 81 View in CoL .
Remark. This name was proposed to be replaced by Barnardetia Xing & Chen, 2013: 399 .
1964 Perinetia Descamps View in CoL : 203, 206 [ Insecta: Orthoptera : Eumastacidae ]; type species: Perinetia annulipes Descamps, 1964: 208 , figs 349–351; by original designation and monotypy.
Remark. This name was proposed to be replaced by Perinetella Descamps & Wintrebert, 1965: 96 . However, this name was preoccupied by Perinetella Synave, 1956: 211 ; type species: Perinetella nigroflava Synave, 1956: 2012 , figs 9–10; by original designation and monotypy ( Insecta: Hemiptera : Flatidae ), and a new replacement name was proposed – Descampsiella Özdikmen, 2008: 67 .
1988 Perinetia Domergue : 135 [ Reptilia: Serpentes: Colubridae ]; type species: Perinetia coulangesi Domergue, 1988 ; by original designation and monotypy.
Remark. This name was proposed to be replaced by Brygophis Domergue & Bour, 1989: 805 .







| NHRS |
Sweden, Stockholm, Naturhistoriska riksmuseet |
| MNHN |
France, Paris, Museum National d'Histoire Naturelle |
| ZIN |
Russia, St. Petersburg, Russian Academy of Sciences, Zoological Institute |
| MMBC |
Czech Republic, Brno, Moravske Muzeum [Moravian Museum] |
| MRAC |
Belgium, Tervuren, Musee Royal de l'Afrique Centrale |
| NHRS |
Swedish Museum of Natural History, Entomology Collections |
| MNHN |
Museum National d'Histoire Naturelle |
| ZIN |
Russian Academy of Sciences, Zoological Institute, Zoological Museum |
| MMBC |
Moravske Muzeum [Moravian Museum] |
| MRAC |
Musée Royal de l’Afrique Centrale |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Laberia palliata Stål, 1866
| Stroiński, Adam, Bourgoin, Thierry & Szwedo, Jacek 2022 |
Aylaella reticulata
| Demir E. & Ozdikmen H. 2009: 271 |
Perinetia
| Descamps M. 1964: 208 |
Perinetia
| Barnard K. H. 1958: 81 |
Perinetia reticulata
| Lallemand V. & Synave H. 1954: 82 |
Perinetia
| Lallemand V. & Synave H. 1954: 82 |
Perinetia
| Seyrig A. 1952: 194 |
Perinetia
| Collenette C. L. 1936: 166 |