
Isometrus longitelson, Deshpande & Gowande & Bastawade & Sulakhe, 2022
|
publication ID |
https://doi.org/ 10.5281/zenodo.7163239 |
|
publication LSID |
lsid:zoobank.org:pub:8B9DDB23-F917-40BD-AEB5-EED78870A7DE |
|
persistent identifier |
https://treatment.plazi.org/id/03EB4B01-7713-FF8A-42DE-A77C0DDBF80F |
|
treatment provided by |
Felipe |
|
scientific name |
Isometrus longitelson |
| status |
sp. nov. |
Isometrus longitelson View in CoL sp. n.
( Figures 1–33 View Figures 1–2 View Figures 3–7 View Figures 8–11 View Figures 12–17 View Figures 18–22 View Figures 23–26 View Figures 27–29 View Figures 30–31 View Figure 32 View Figure 33 , Tables 1, 2) http://zoobank.org/urn:lsid:zoobank.org:act:B6C82C3A-
BF0E-4D96-BD65-782FC266DEDC
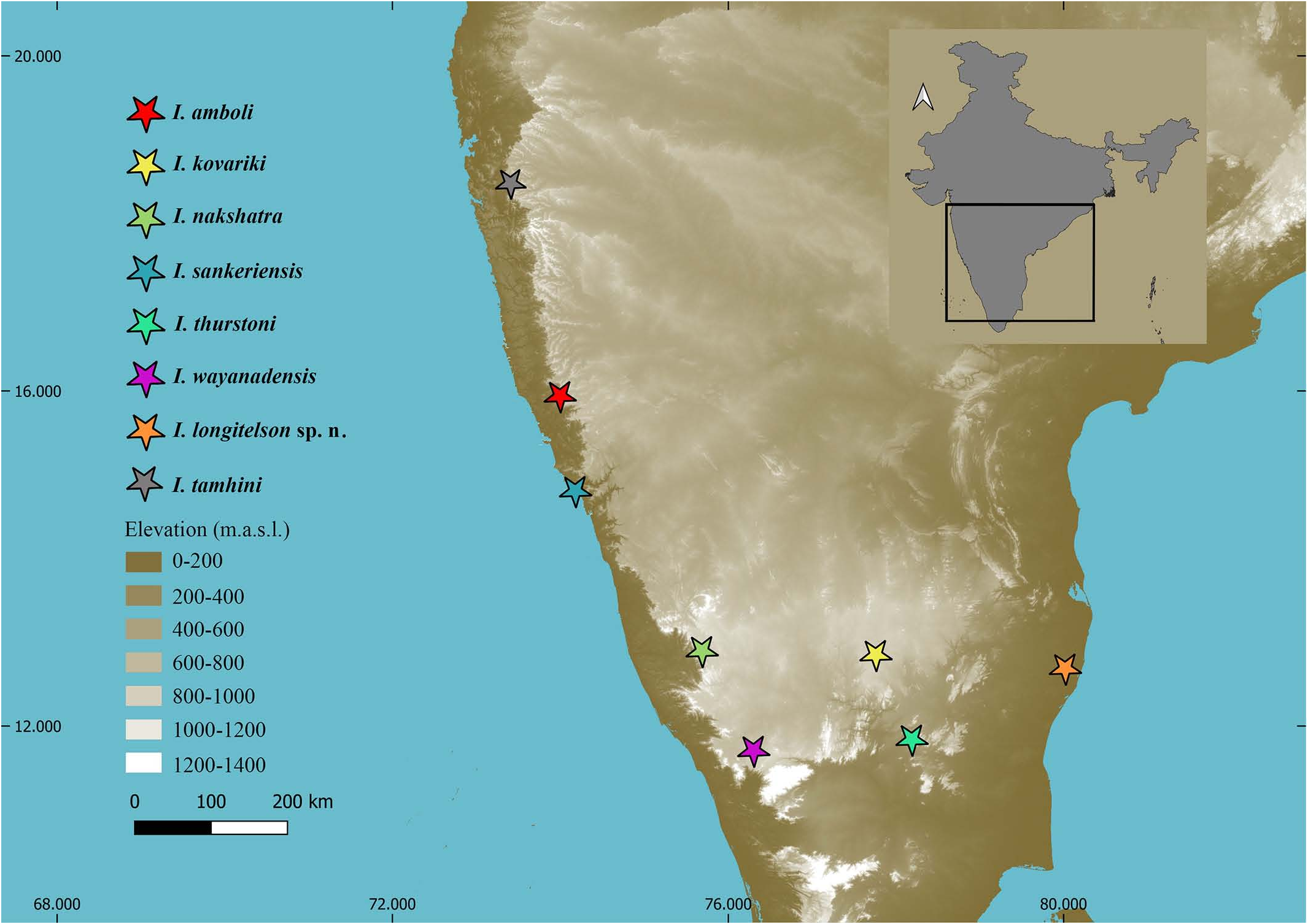
TYPE LOCALITY AND TYPE REPOSITORY. India, Tamil Nadu State, Chengalpattu District, Keelavedu Village, ChengalpattuThiruporur Road , 12.68˚ N 80.01 ˚E, 57 m a. s. l. ; BNHS.
TYPE MATERIAL. India, Tamil Nadu State, Chengalpattu District, Keelavedu Village, Chengalpattu-Thiruporur Road , 12.68˚ N 80.01 ˚E, 57 m a. s. l., leg. S. Datta, 27 September 2016, 1 ♂ (holotype, BNHS SC 287 View Materials ) 4♂ (paratypes, IES 368 , 370 , BNHS SC 288 View Materials , 289 View Materials ) 4♀ (paratypes IES 366 , 369 , 372 , BNHS SC 290 View Materials ) .
ETYMOLOGY. The specific epithet is an adjective referring to the elongated telson in the males of the species
DIAGNOSIS. (♂ ♀) Total length 37.10–54.72 mm. Base colouration yellowish-brown and variegated with blackbrown stripes and spots. Basal segments of chelicerae dorsally yellowish with blackish reticulation. Pectinal tooth number 15–18 in both sexes. Median supra-ocular area, finely granular. Median ocelli anteriorly situated in the ratio 1:2.2. Promedian carina on pedipalp patella strongly developed. Tergites I–VI finely granular with median carina more strongly developed on posterior side.
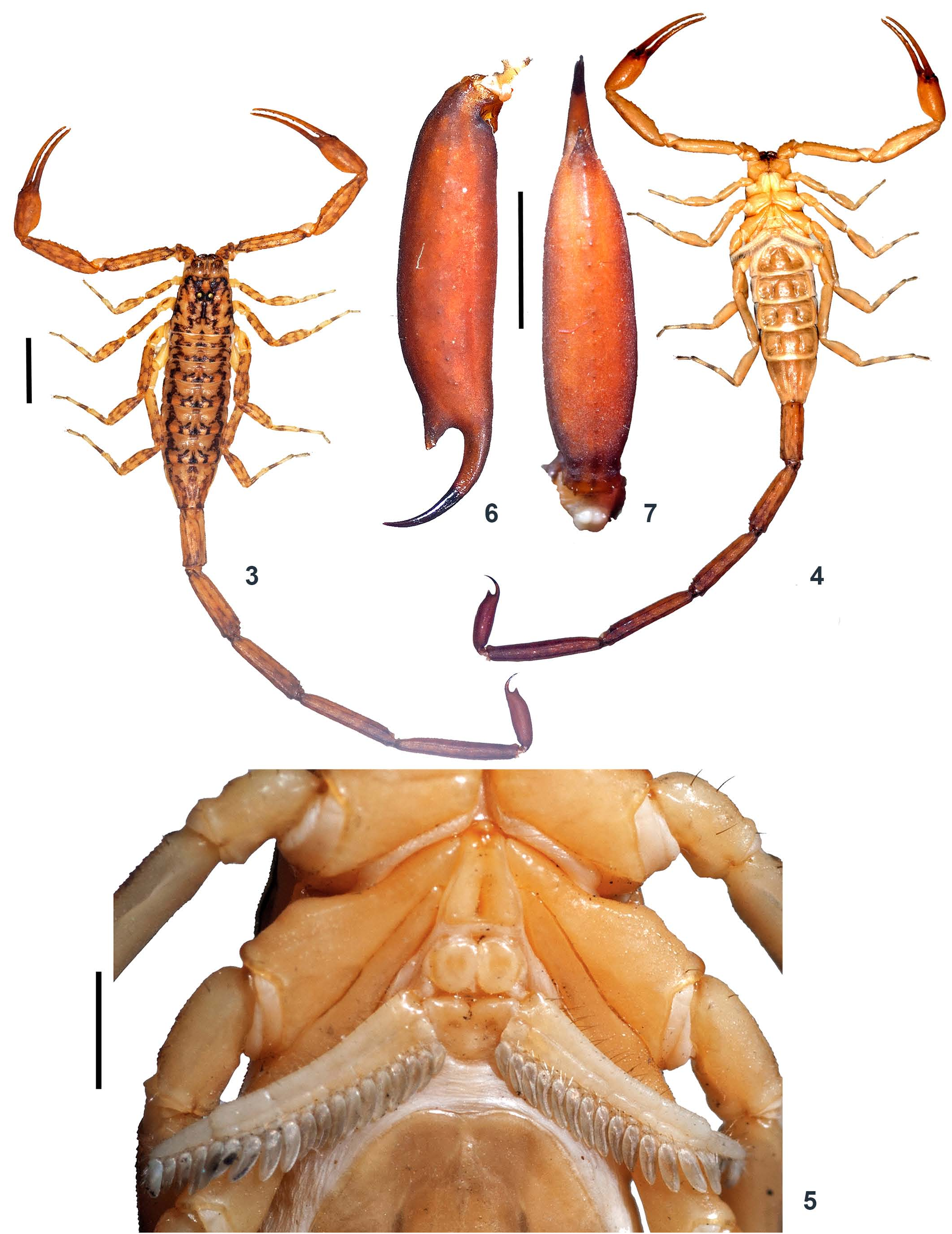
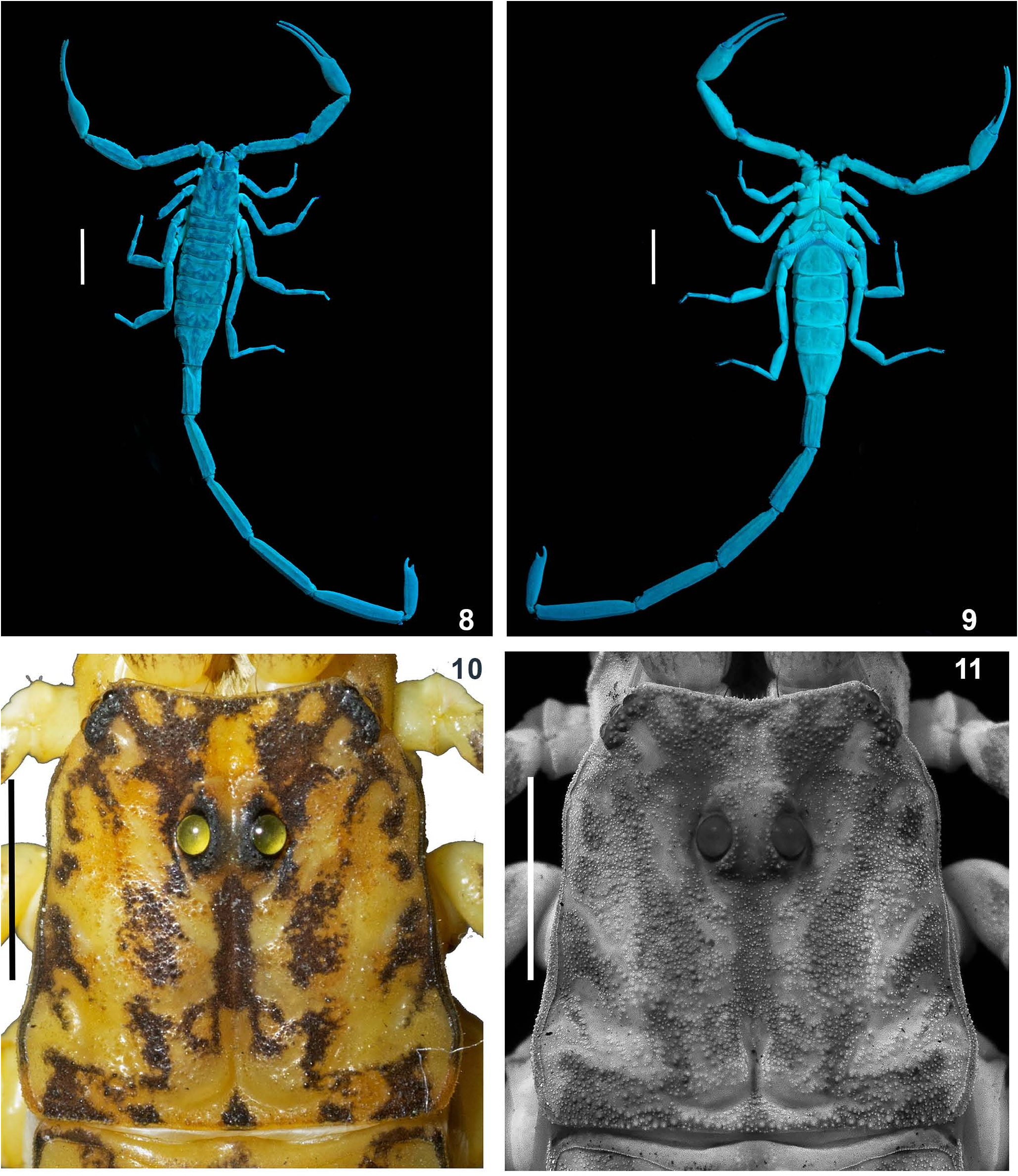
DESCRIPTION (♂ holotype; measurements in Table 1) Coloration ( Figs. 3–5 View Figures 3–7 , 10 View Figures 8–11 ). Body yellowish brown and variegated with blackish brown stripes and spots; appendages yellowish with blackish brown stripes and spots; metasomal segment V dark brownish to blackish, darker on posterior portion; pedipalp fingers yellowish with dark brown on one third proximal portion. Ventral portion uniformly yellow. Basal segments of chelicera yellow with blackish reticulation ending anteriorly into blackish transverse thin patch. Fingers of chelicera dark brown with tip of the fingers black. Telson dark brown in color.
Carapace ( Figs. 10, 11 View Figures 8–11 ). Surface densely granular with coarse and fine granules (mixed). Carapace without carinae. Median supra-ocular area, finely granular. Inter-ocular area with fine and dense granules. A pair of median eyes situated anteriorly in the ratio 1:2.2 (ratio of median eyes to anterior margin and median eyes to posterior margin). Antero-lateral ocular tubercle granular with type 5 lateral ocelli. Three pairs of large major ocelli and two small minor ocelli situated behind the major ocelli. Longitudinal furrow shallow anteriorly and deep posteriorly. Anterior margins finely granular. Lateral margins weakly crenulated below lateral ocelli. Posterior margin almost entirely smooth.
Chelicerae. Characteristic of the family Buthidae . Basal segments and movable fingers with tuft of short and firm setae on basal and ventral surfaces.
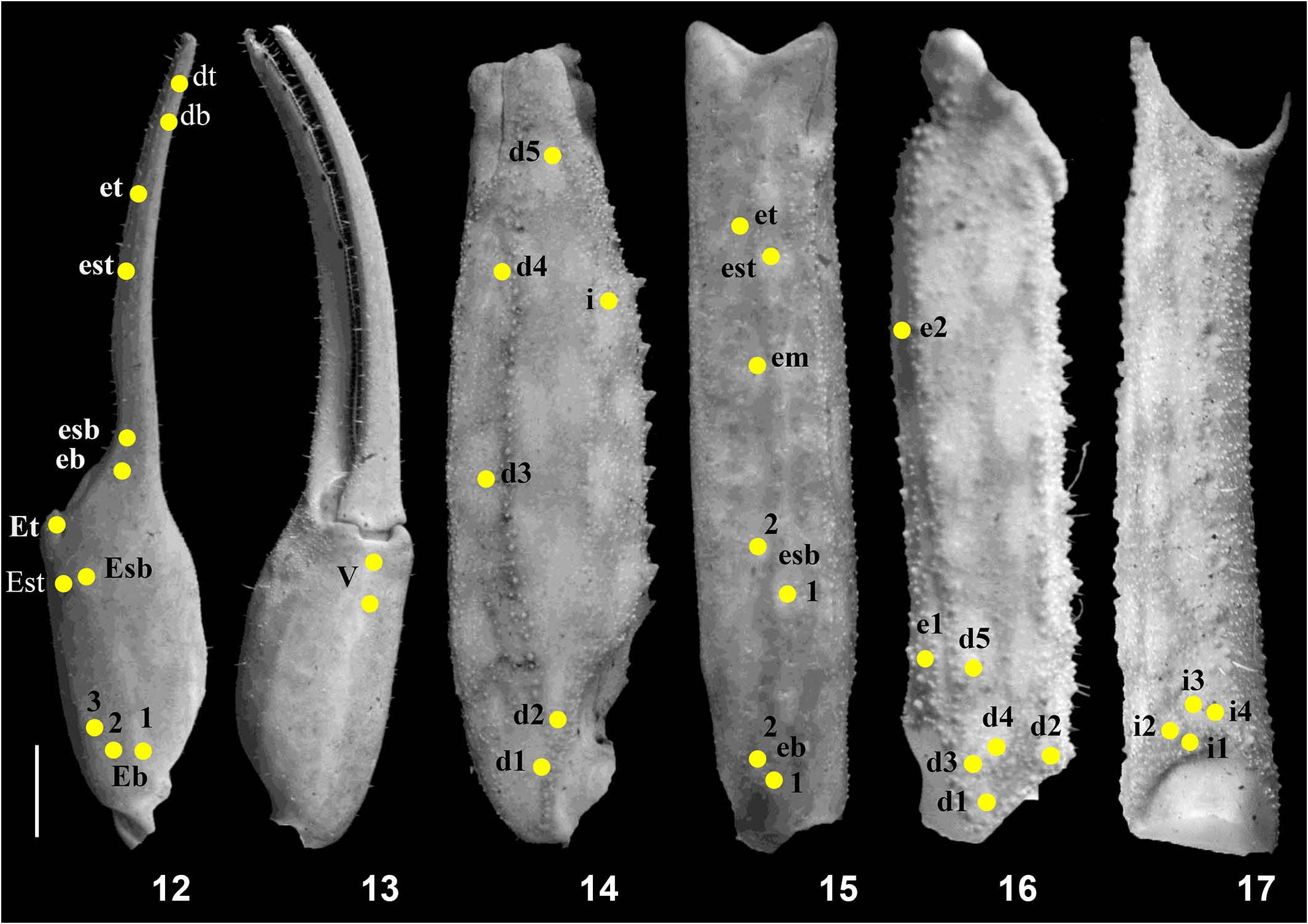
Pedipalp ( Figs. 12–17 View Figures 12–17 ). Femur with five carinae (prodorsal, retrodorsal, promedian, retromedian and proventral). All carinae crenulated. Intercarinal surfaces with mixed and sparse granulation except ventral surface smooth with few fine granules on proximal portions. Patella with seven distinct carinae (dorsomedian, prodorsal, retrodorsal, retromedian, retroventral, promedian and proventral). Intercarinal surfaces weakly granular on dorsal surface and smooth on ventral surface. Manus with four carinae (dorsomedian and dorsoretrosubmedian accessory prominent; retromedian and retroventral obsolete). Fixed fingers with two smooth digital and inner secondary carinae. Movable and fixed fingers slightly scalloped with six rows of prolateral and retrolateral denticles in pairs and one additional single row of retrolateral denticles on proximal portion. Trichobothrial pattern of type A, typical for the genus (chela dorsal 12, chela ventral 2, patella dorsal 6, patella retrolateral 7, femur dorsal 7 and femur prolateral 4). Legs ( Figs. 3, 4 View Figures 3–7 , 8, 9 View Figures 8–11 ). Femur and patellae carinated. All anterior carinae granular whereas all posterior carinae smooth. Tibia 3 and 4 without tibial spur. All legs with a pair of pedal spurs. Tarsomere covered with long delicate setae arranged in parallel rows on ventral side. Tarsomere I with a tuft of short, stout blackish setae on ventral side. Tarsomere II compressed laterally and ventrally with paired row of short, pointed, anteriorly directed, closely placed setae.
Genital operculum ( Fig. 5 View Figures 3–7 ). Wider than long, elliptical, separated with a pair of short male genital papillae.
Pectines ( Fig. 5 View Figures 3–7 ). Basal piece rectangular, notched on anterior median margin. Posterior margin of basal piece curved. Marginal lamella of 3/3 digits and median lamella of 6/7 digits, outer margin armed with a row of stout short red setae and few setae on surface. Fulcra 17/18, very small, roughly triangular, each armed with few short red setae, placed in between adjacent pectinal teeth. Teeth 17/18, strong and stout. Mesosoma ( Figs. 3, 4 View Figures 3–7 , 8, 9 View Figures 8–11 , 26 View Figures 23–26 ). Tergites I–VI finely granular with median carina more strongly developed on posterior side. Posterior and lateral margins granular. Coarse granulation on mesosomal tergites V and VI along the margins. Tergite VII granular, narrowed posteriorly, with two pairs of lateral granular carinae. Broad median carina present, more strongly developed on anterior portion. Sternites III–V almost entirely smooth, each with a pair of spiracles. Sternite VI finely granular on lateral portion. Sternite VII entirely granular, more closely granular on lateral portion; two pairs of granular carinae present on two third posterior portion.
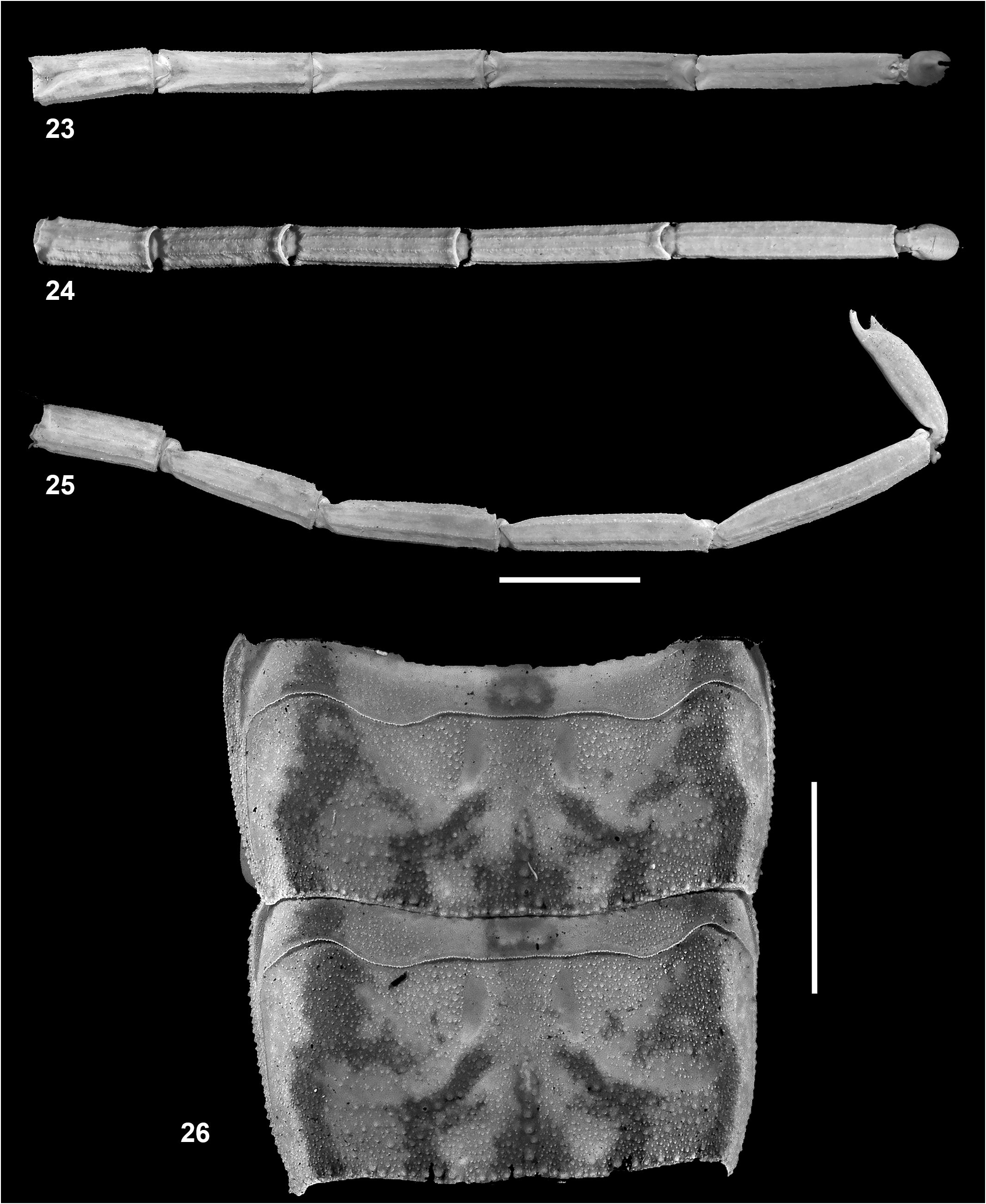
Metasoma ( Figs. 3, 4 View Figures 3–7 , 8, 9 View Figures 8–11 , 23–25 View Figures 23–26 ). All segments longer than wide. Segment I with five pairs of granular carinae (dorsal lateral, lateral supramedian, lateral inframedian, ventral lateral and ventral submedian). Segments II–IV with four pairs of carinae (dorsal lateral, lateral supramedian, ventral lateral and ventral submedian). Lateral supramedian and ventral lateral carinae on segments II and III moderately granular. Intercarinal surfaces almost smooth. Dorsal lateral carina on segments I–IV ending in very weak tubercles. Segment V with five carinae (dorsal lateral, lateral supramedian, ventral lateral pairs, ventral submedian and single ventral median). Dorsal lateral carina weakly developed. Intercarinal surfaces almost smooth. Anal rim weakly granular.
Telson ( Figs. 2 View Figures 1–2 , 6–9 View Figures 3–7 View Figures 8–11 , 25 View Figures 23–26 ). Telson with slender and elongated vesicle, smooth on dorsal surface. Ventral median carina weakly granular, ending in triangular, subaculear, pointed nodule, armed with a pair of minute denticles on inner basal margin. Lateral and ventral intercarinal surfaces weakly granular. Aculeus elongated and moderately curved.
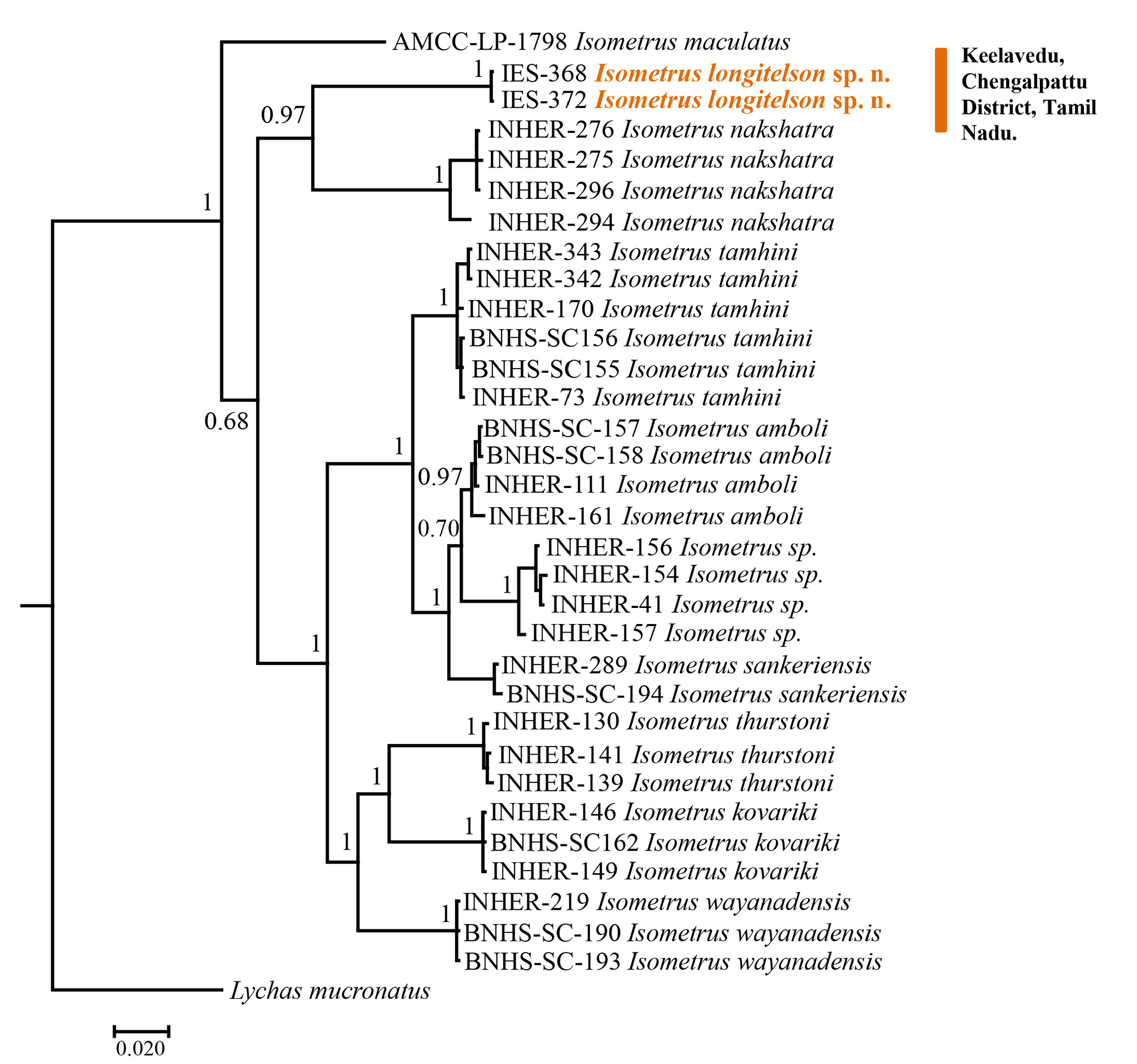
DISTRIBUTION AND ECOLOGY. I. longitelson sp. n. is only known from its type locality, Chengalpattu-Thiruporur Road, Keelavedu Village, Chengalpattu District, Tamil Nadu, India. All the specimens were collected from the tree bark of Tamarindus indica Linné, 1753 (Tamarind trees) adjacent to the road just after dusk at an elevation of 4–6 feet from the ground. A population of the new species was observed on these trees along the entire road. Multiple individuals were observed on single trees. The new species was observed to be strictly arboreal living below the tree bark similar to all the other species of Isometrus found in India. The new species might be occurring in the forest on the hills surrounding the Chengalpattu-Thiruporur Road, however this needs to be confirmed. It is interesting to note that the new species was observed to be closely associated with the Tamarind trees present in the type locality. The precise origin of Tamarindus indica is still subject to controversy. It is claimed ( Lefévre 1971; Grollier et al., 1998; El-Siddig et al., 1999) that it originated in Africa and was only introduced into India; however, Wunderlin (1998) and Chauvin & Poupon (1983) assumed that its origin is in Asia, particularly in India, because of its appellation “Tamar hindi”, which means “Indian date”. This close association between the two species might support the later hypothesis; however this needs to be confirmed with biogeographic analysis ( Figs. 27–29 View Figures 27–29 , 33 View Figure 33 ).
AFFINITIES. Isometrus longitelson sp. n. differs from all other Indian species of Isometrus based on the following set of morphological characters:
1. Surface of carapace with mixed (coarse and fine) and dense granulation ( Figs 10, 11 View Figures 8–11 ) as opposed to coarse and dense granulation in I. tamhini and I. wayanadensis ; coarse and sparse granulation with some areas without granules in I. sankeriensis and I. thurstoni ; granular throughout with mixed granules, more densely granular in inter-ocular area and median posterior ocular area in I. kovariki and granular throughout but obsolete in I. maculatus .
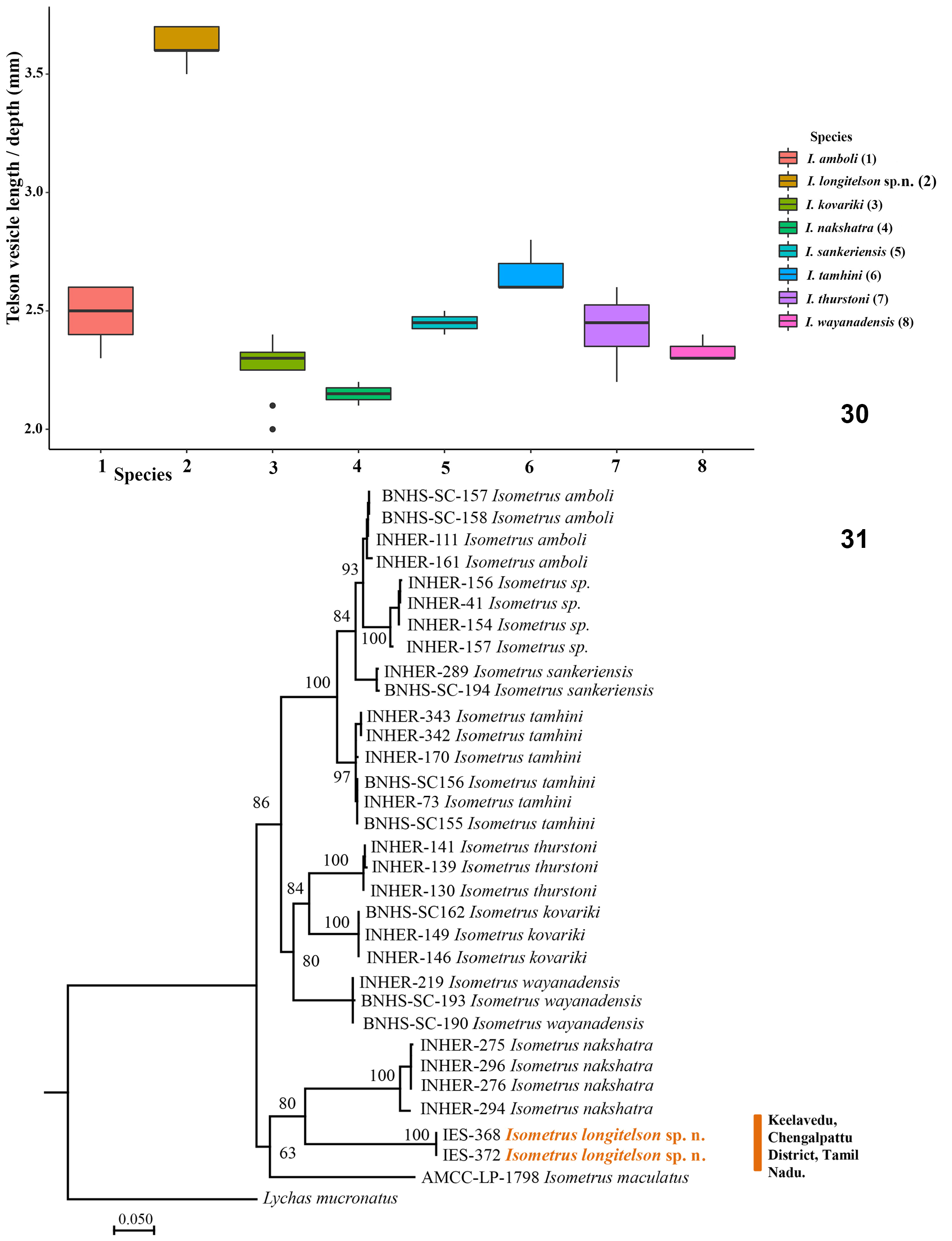
2. Telson vesicle length to depth ratio in males 3.5–3.7 as opposed to 2.6–2.8 in I. tamhini ; 2.3–2.6 in I. amboli ; 2.4–2.5 in I. sankeriensis ; 2.2–2.6 in I. thurstoni ; 2.0– 2.4 in I. kovariki ; 2.1–2.2 in I. nakshatra and 2.3–2.4 in I. wayanadensis ( Figs. 1 View Figures 1–2 , 6, 7–9 View Figures 3–7 View Figures 8–11 ).
3. Coarse granulation on mesosomal tergites V and VI along the margins ( Fig. 26 View Figures 23–26 ) as opposed to fine granulation along margins in I. wayanadensis , I. nakshatra , I. sankeriensis , I. amboli , I. thurstoni and I. kovariki .
4. Chela length to width ratio in males 5.0 – 5.4 as opposed to 6.1–6.5 in I. tamhini , 5.7–5.8 in I. sankeriensis and 10.6 in I. nakshatra and in females 5.0–5.4 as opposed to 5.7 in I. amboli , 4.8 in I. kovariki and 6.1–6.2 in I. nakshatra ( Table 1). 5. Metasomal length to carapace length ratio in males 8.2–8.9 as opposed to 5.9–6.1 in I. sankeriensis , 6.5–7.3 in I. kovariki , 5.3–5.8 in I. nakshatra , 6.8–7.2 in I. wayanadensis 9.6 in I. maculatus ( Table 1).
6. Lateral supramedian and ventral lateral carinae on metasomal segments II–IV moderately to weakly granular ( Fig. 23–25 View Figures 23–26 ) as opposed to strongly granular in I. tamhini and I. sankeriensis .
7. Ventral median carina of telson vesicle weakly granular ( Fig. 7 View Figures 3–7 ) as opposed to moderately granular in I. amboli , I. wayanadensis and strongly granular in I. tamhini .
8. Spiniform granules of promedian carina of the pedipalp patella strongly developed ( Fig. 14 View Figures 12–17 ) as opposed to moderately developed in I. tamhini , I. amboli , I. sankeriensis , I. kovariki and weakly developed in I. nakshatra .



| BNHS |
Bombay Natural History Society |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.