
Ischyropsalis cantabrica, Luque, Carlos G. & Labrada, Lucia, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.211031 |
|
DOI |
https://doi.org/10.5281/zenodo.6179950 |
|
persistent identifier |
https://treatment.plazi.org/id/84708794-EF45-FFA6-FF70-9B726748F8FF |
|
treatment provided by |
Plazi |
|
scientific name |
Ischyropsalis cantabrica |
| status |
sp. nov. |
Ischyropsalis cantabrica View in CoL sp. n.
( Figs 1–24 View FIGURES 1 – 3 View FIGURE 4 View FIGURE 5 View FIGURES 6 – 10 View FIGURES 11 – 15 View FIGURES 16 – 17 View FIGURES 18 – 21 View FIGURES 22 – 24 )
Type material: Holotype 3 ( MNCN 20.02/12828) and Allotype Ƥ ( MNCN 20.02/12829): SPAIN, Cantabria: Alfoz de Lloredo, Oreña: Rogeria Cave (30TVP0867005271, 90 m), 2-IV-1994, leg. C.G. Luque. Paratypes: SPAIN, Cantabria: Alfoz de Lloredo: 5 3, 4 ƤƤ, same data as the holotype (1 3, 1 Ƥ MNCN; 1 3, 1 Ƥ NHMW, 1 3, 1 Ƥ AXLS, the rest CGL); Ruiloba, Casasola: 1 3, 1 Ƥ, Portillo Cave (30TVP9639204775, 20 m), 25-III-1995, leg. C.G. Luque & E. Muñoz (CGL); Cabezón de la Sal, Bustablado: 1 3 mounted on SEM stubs ( MNCN 20.02/12857), Cerviz Cave (30TVP0286800238, 365 m), 1-IV-1995, leg. C.G. Luque & J. Ruiz; Alfoz de Lloredo, Oreña: 2 3, Calero Cave, 12-VIII-2003, leg. L. Labrada & C.G. Luque (CGL).
Additional material studied: SPAIN, Cantabria: Udías, Canales: 1 Ƥ ( MNCN 20.02/12863), Cáscaras Cave (30TVP 9894201878, 130 m), 2-IX-1915, leg. C. Bolívar (see Bolívar 1916, p. 319).
Diagnosis: A species of the kollari group (sensu Martens 1969) with long and slender chelicerae (spines completely absent in males); cheliceral apophysis high, not projected forwards, with rounded conical shape and the piliferous apex, with a series of deep parallel grooves, medial and characterized by an additional small piliferous and pocket-forming apophysis; dorsal region of the glans penis with a dense mass of very long bristles at its end; glans short and conical shaped at its apex; upper part of glans rounded-quadrangular shaped in cross-section; sclerite of glans long and narrow, almost parallel-sided, basal end tapering, median keel developed, bristle cover dense, widely separated into two lateral areas; body depigmented but scutum parvum uniformly dark colored (dorsal abdominal sclerites fused).
Description:
Measurements were taken for the holotype, allotype and paratypes (9 3 and 6 ƤƤ). Body length (with distended abdomen): holotype 5.01, 3 paratypes 5.01–5.45, allotype 5.58, ƤƤ paratypes 5.56–5.81; length of basal cheliceral segment (in parentheses, with length/width of distal cheliceral segment): holotype 5.20 (6.50/0.80), 3 paratypes 5.20–5.54 (6.50–6.88/0.80), allotype 5.53 (7.25/0.84), ƤƤ paratypes 5.53–5.85 (7.25–7.68/0.84). Female show a wider range in body length due to different stages of gravidity but their chelicerae are less variable in size than in male. Total length of leg II of male holotype (paratypes in parentheses) and female allotype (paratypes in parentheses): 42.49 (42.49–44.82), 42.57 (42.57–43.78). Length of the II segments (in parentheses as before): femur: 8.82 (8.82–8.98), 9.32 (9.32–9.66); patella: 1.36 (1.36–1.41), 1.49 (1.49–1.56); tibia: 8.38 (8.38–8.94), 8.41 (8.41–8.78); metatarsus: 13.52 (13.52–13.89), 13.40 (13.40–13.81); tarsus: 10.41 (10.41–10.82), 9.95 (9.95–10.21). The measurements of the male (holotype) and female (in parentheses) femurs from the first to the fourth pair of legs are 6.45 (6.84), 8.82 (9.32), 5.34 (5.56), 7.08 (7.35).
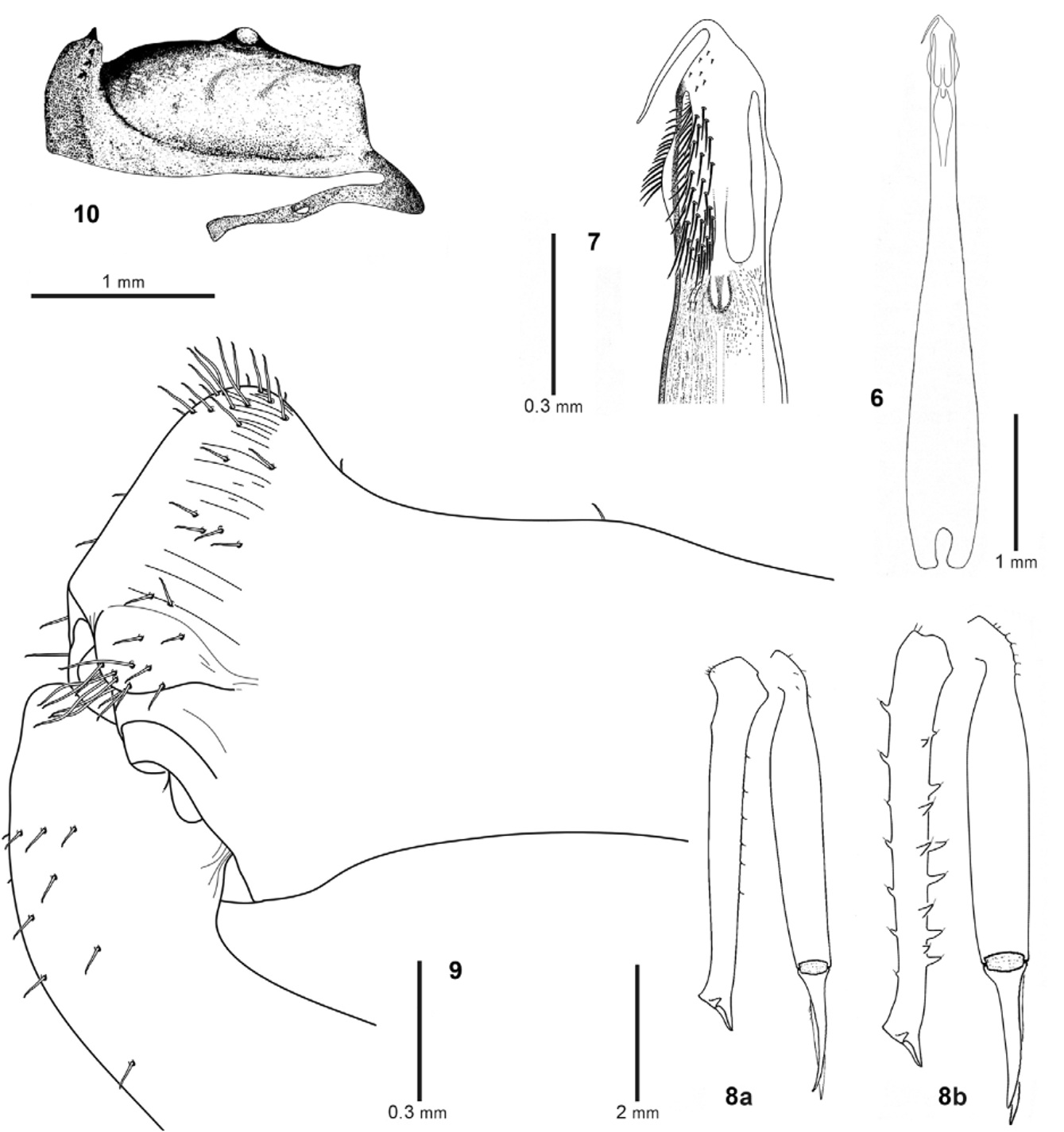
Genital morphology ( Figs 6–7 View FIGURES 6 – 10 , 11–15 View FIGURES 11 – 15 ). Penis: Symmetrical, with robust truncus and the base long and split ( Fig. 6 View FIGURES 6 – 10 ); the truncus is of medium length (L: 4.0). The coloration is dark, from the truncus to the base, the ventral side is depigmented; the glans in contrast is blackish in colour. The glans at its apex is short conical shaped, with a long stylus ( Figs 7 View FIGURES 6 – 10 , 13 View FIGURES 11 – 15 ). On the dorsal side ( Fig 11 View FIGURES 11 – 15 ), the glans has a dense mass of very long bristles at its ends (the bristles become shorter nearer the apex of the glans). In the upper part of glans, the outline of the cross section is rounded-quadrangular shaped ( Fig 11 View FIGURES 11 – 15 ) and does not narrow gradually to acquire a triangular shape in lateral view. In contrast, the ventral side displays a sclerite of glans long and narrow, almost parallel-sided, basal end tapering, ventral median keel developed, bristle cover dense, widely separated into two lateral areas ( Fig 12 View FIGURES 11 – 15 ), and a large depigmented mouth indicating the attached muscular tendon ( Fig 15 View FIGURES 11 – 15 ). The glans is clearly inflated in lateral view, appearing swollen in mid-section. This spaces the ventro-lateral bristle areas wide apart from each other. The midarea has a thin longitudinal row of short bristles ( Figs 13–14 View FIGURES 11 – 15 ). Ovipositor: bilobulate, with the ventral apex covered with few hairs; the seminal receptacle has four short tubes. In this respect not differentiated from other Ischyropsalis species ( Martens 1969; Dresco 1968a, p. 313, fig. 19; Dresco 1968b, p. 966, fig. 20; Dresco 1972b, p. 356, fig. 25; see also Gruber 2007, p. 147, fig. 4.15i).
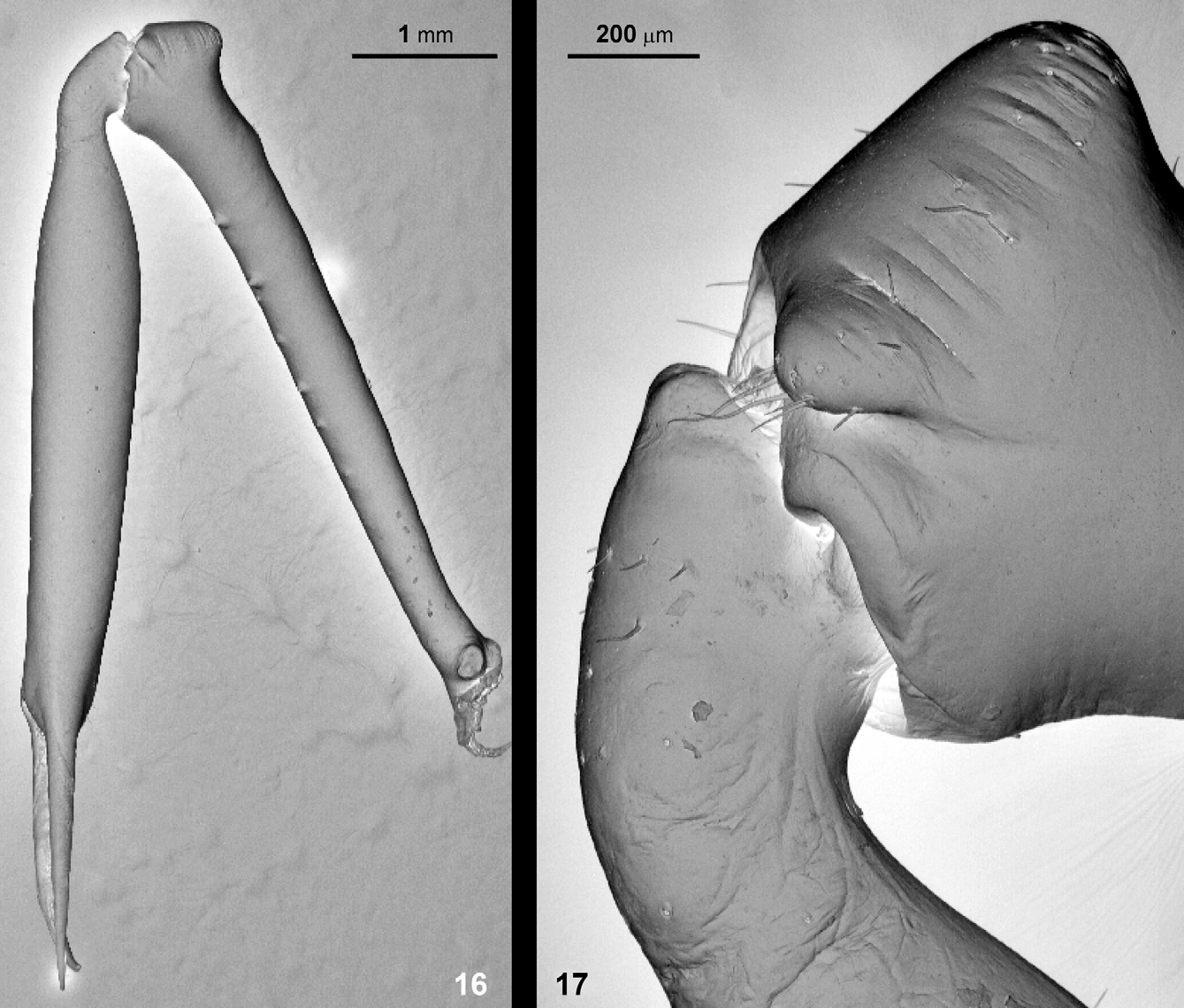
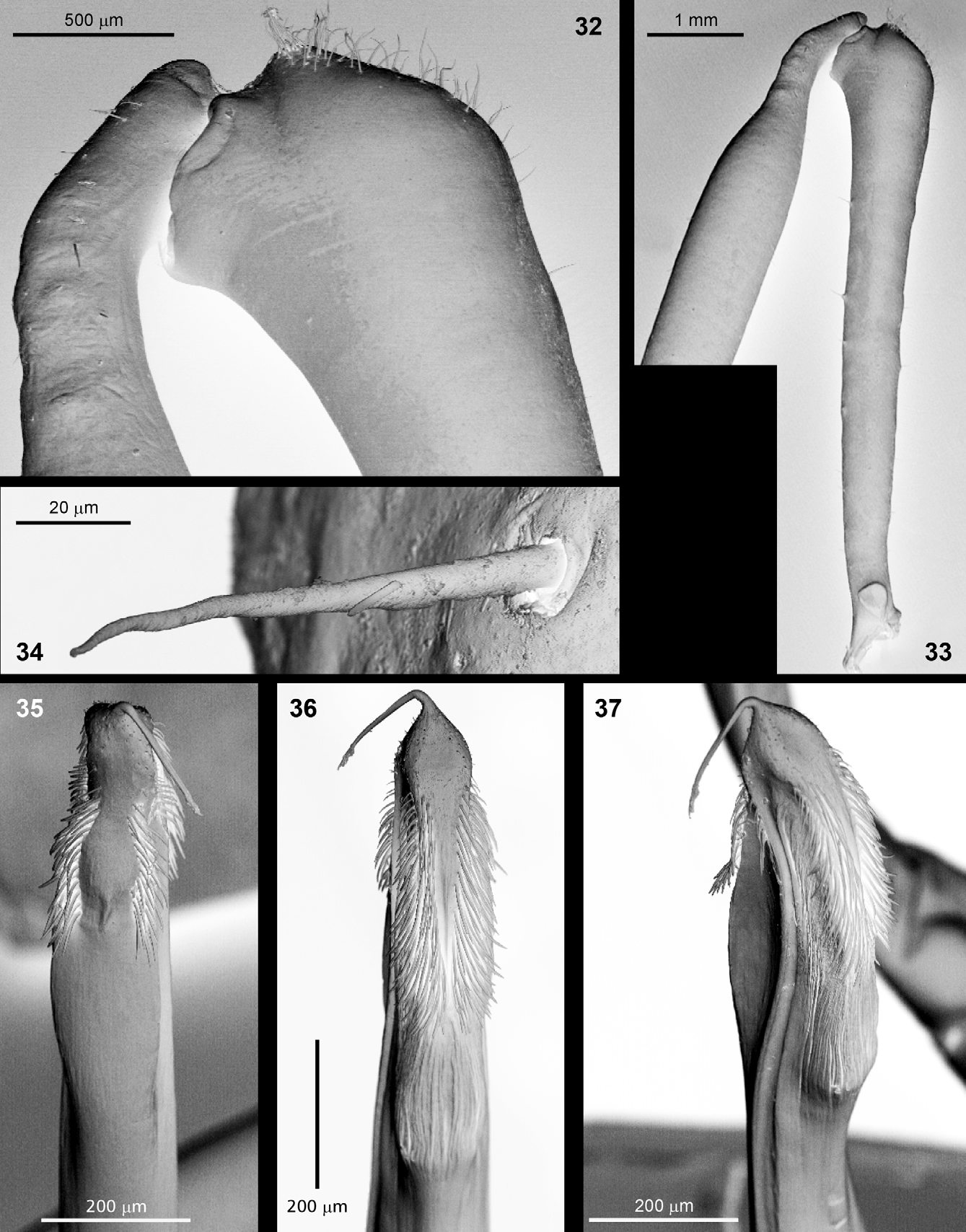
Chelicerae ( Figs 8–9 View FIGURES 6 – 10 , 16–17 View FIGURES 16 – 17 ): Sexually dimorph. In general, long and slender, black or dark brown. In males basal segment with a longitudinal row of small and flat spines with bristles ventrally and with 4 or 5 piliferous tubercles dorsally ( Figs 8 View FIGURES 6 – 10 a, 9). Basal segment narrow at the base, widens distally to a pronounced triangular apophysis (lateral view; Figs 8 View FIGURES 6 – 10 a, 9, 16–17). The apophysis does not project proximal but dorsal, and has a rounded conical shape with the piliferous apex, with a piliferous pocket-forming apophysis and a series of deep parallel grooves on the inner side ( Figs 9 View FIGURES 6 – 10 , 17 View FIGURES 16 – 17 ). The distal segment is completely smooth, only few short hairs present at the base ( Figs 8 View FIGURES 6 – 10 , 16–17 View FIGURES 16 – 17 ). They are generally bristle-like and are set into a basal socket ( Fig 34 View FIGURES 32 – 37 ). The chelicerae of females are larger, with basal segment more robust and armed with long spines of different size over its whole length (six shorter spines on dorsal surface, ventro-medially with six, and ventro-laterally with five spines; Fig 8 View FIGURES 6 – 10 b). The distal segment is covered with small rounded piliferous tubercles at the dorsal knee, and also with some short hairs towards the base of the pincer.
Prosoma: Cephalothorax flat ( Fig 10 View FIGURES 6 – 10 ), black or dark brown, moderately ascending from border of second thoracic tergite, and ornamented with a transversal row of bristles with six denticles of different sizes; eye mound weakly developed, eyes widely separated, with small lenses in comparison to I. gigantea ( Fig 27 View FIGURES 27 – 31 ).
Opisthosoma: Males and females with scutum parvum, the last three tergites always free. Sclerotised area finely grained, surrounded by slight grooves, each with a transversal line of bristles ( Fig 3 View FIGURES 1 – 3 ).
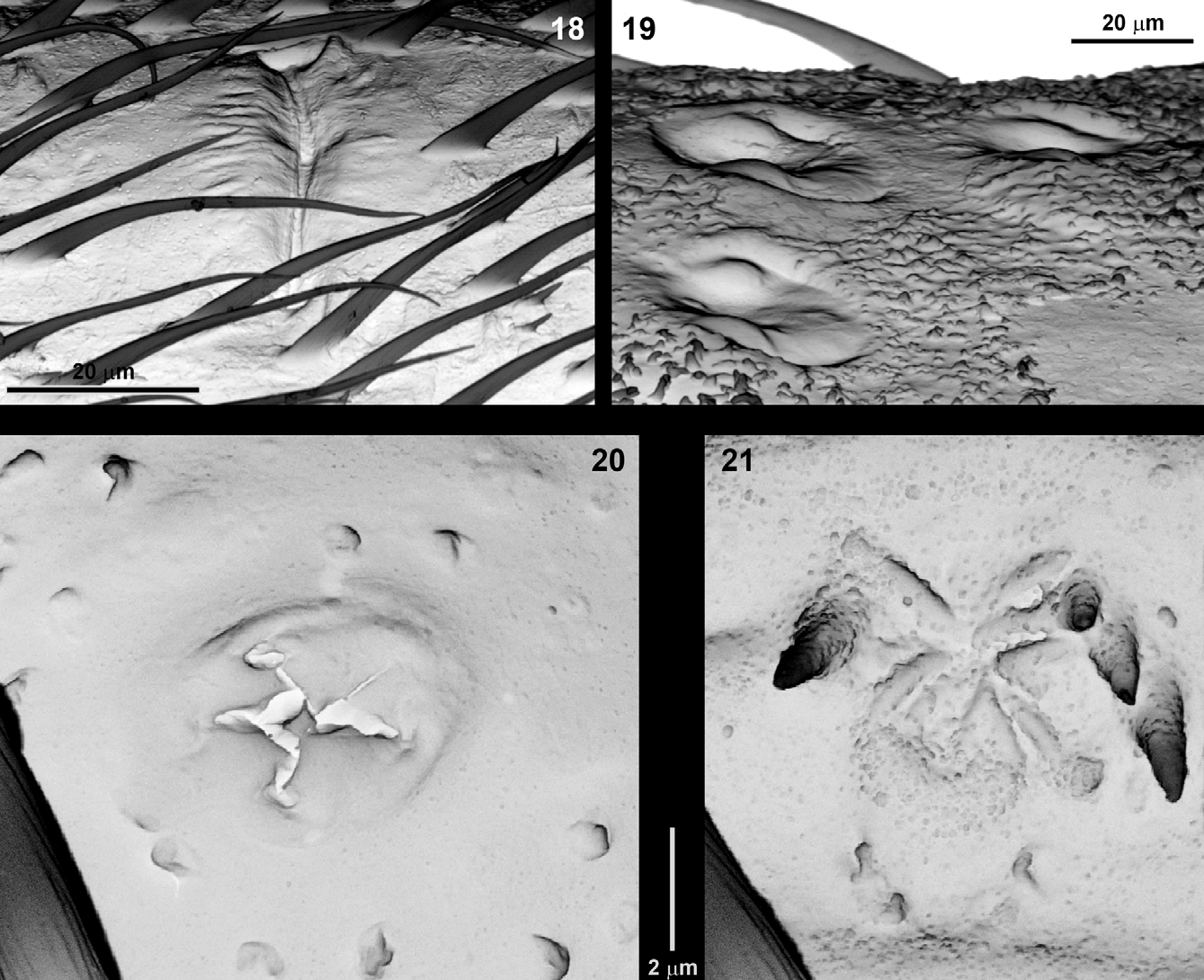
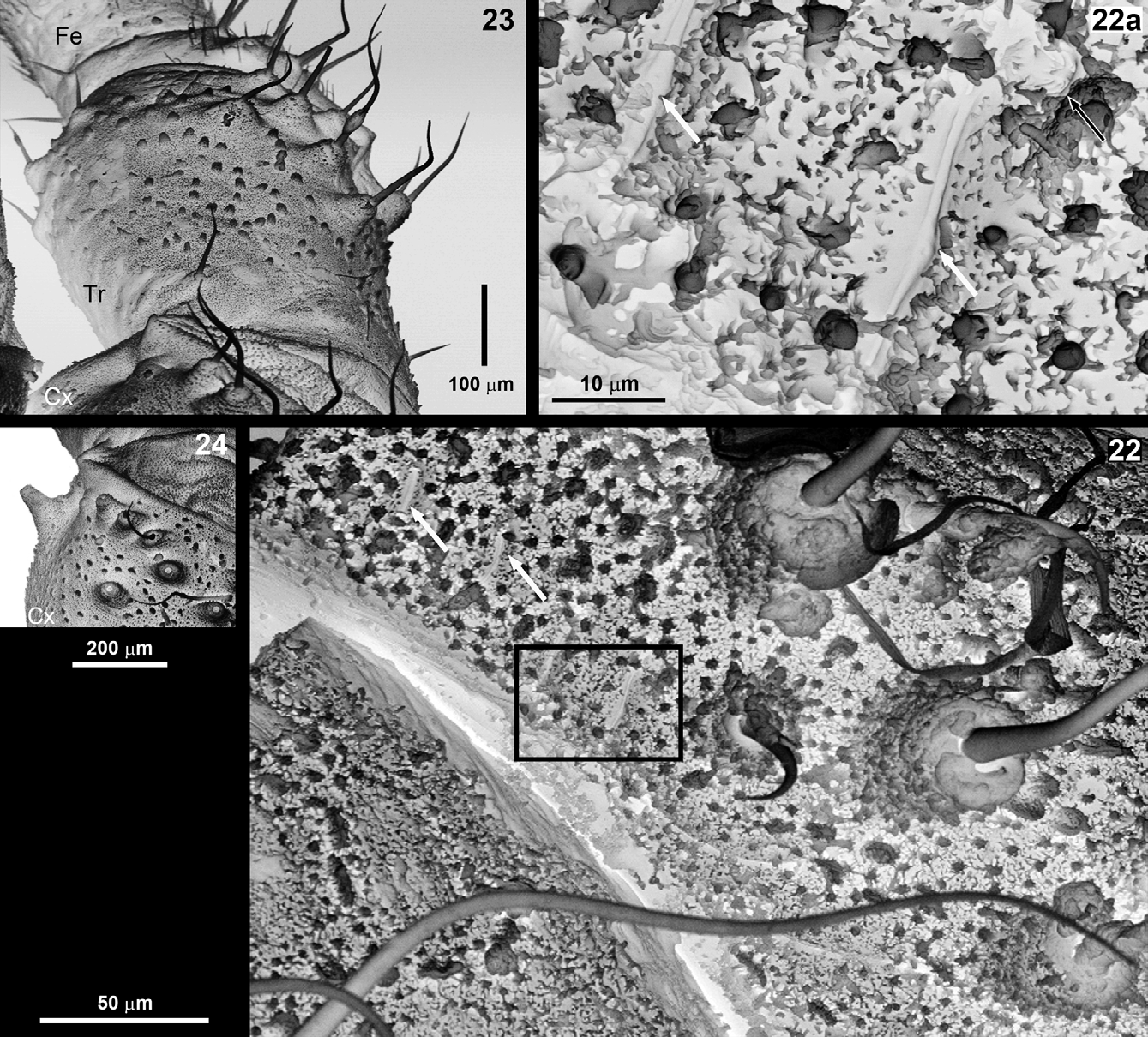
Legs: Slender, of medium length, all segments rounded, light brown, metatarsus and tarsus of lighter colour, base of femora whitish to yellowish; femora without sculpture elements (spines or tubercles), only with microtrichia cover and setae; surface of basal segments (trochanter and coxa) with several types of sculpture elements (e.g. granules, tubercles and cones), with long macrotrichia, which arise from a basal socket ( Figs 18–24 View FIGURES 18 – 21 View FIGURES 22 – 24 ).
Variability: There seems to be no extraordinary variation despite the general differences in Ischyropsalis species, e.g. in case of females spination is slightly more pronounced in larger specimens (see Martens 1969, pp. 160–165). The structure of the glans penis is relatively constant. This is also the case for the other species of the Iberian kollari group (sensu Martens 1969), except for the heterogeneous I. dispar group as shown in Figs 38–41 View FIGURES 38 – 41 and 50–57 View FIGURES 50 – 57 (see discussion).
Etymology: The species is named for the Cantabria Region, for which it is endemic.
Distribution ( Figs 4–5 View FIGURE 4 View FIGURE 5 ): Endemic to the northwestern Cantabrian coastal region in Spain, and restricted to a narrow band of karst systems along the coast within the municipal districts of Alfoz de Lloredo, Ruiloba, Udías and Cabezón de la Sal. Epigean species, like I. nodifera and I. petiginosa , occur in all caves visited along the Cantabria coast (see, e.g., Luque 1991), while restricted to the shallower zone (<30 m) and not interacting with I. cantabrica sp. n.
Ecology: The caves inhabited by I. cantabrica sp. n. developed in the Lower Cretaceous (urgonian-type facies) limestones (Aptian age). All specimens were collected in the lightless zones of caves (especially in “deep cave” sites), about 300 m from the entrance of Rogeria Cave ( Fig. 1 View FIGURES 1 – 3 ), and about 150 m from the entrances of both Portillo and Cerviz Caves. The specimens were usually found close to water, on cave walls with abundant speleothems or mud-cover, and between 0.5 to 2 m above water level. In the Calero Cave I. cantabrica sp. n. was observed to prey on the small aquatic gastropod Ancylus sp. aff. fluviatilis (in spring of late to mid-1970s; Menéndez 1973). Although they are mainly active predators, in some cases they do not disdain carrion. Sixteen other invertebrates were recorded from the deep cave zone of all the caves inhabited by I. cantabrica sp. n. ( Menéndez 1973; Labrada & Luque 2004): Trichoniscoides cavernicola (class Crustacea, order Isopoda ); Lithobius validus and L. tricuspis (both class Chilopoda, order Lithobiomorpha ); Polydesmus coriaceus , Trachysphaera rousseti and Blaniulus dollfusi (all three class Diplopoda, orders Polydesmida , Glomerida and Iulida); Litocampa espanoli (class Insecta, order Diplura ); Pseudosinella superoculata and Arrhopalites sp. aff. elegans (both class Insecta, order Collembola ); Neobisium (Neobisium) sp. aff. cavernarum (class Arachnida, order Pseudoscorpionida); Nemastoma (s. lat.) sexmucronatum (class Arachnida, order Opiliones ); Troglohyphantes cantabricus and Nesticus luquei (both class Arachnida, order Araneae ); Quaestus (Quaestus) arcanus and Laemostenus (Antisphodrus) pelaeus (both class Insect, order Coleoptera ).
Conservation: Although I. cantabrica sp. n. seems frequent in suitable habitats, which are numerous throughout its area, it is an extremely narrow endemic species and therefore requires particular conservational attention. So far, the species is not listed as ‘vulnerable’ under the Threatened Species Act of 2008 (enacted under Decree No. 120/2008, and regionally known as ‘Catalogue of Threatened Species from Cantabria’) or the Nature Conservation Act of 2006 (under the Cantabria Regional Government legislation; Labrada et al. 2010). These legislations cover the protection of a wide range of species (e.g. cave-dwelling bats and invertebrates such as cavedwelling beetles Quaestus arcanus ) and habitats (e.g., in the case of Rogeria Cave). This cave is further acknowledged as ‘important bat underground habitat’ ( Menéndez 1973; Labrada & Luque 2004, unpublished results). Access to the cave is currently restricted, and occasionally prohibited, to protect threatened cave-dwelling bats —e.g. Rhinolophus euryale , Miniopterus schreibersii and Myotis myotis — particularly during their breeding season (from June to August-September). For this reason, Rogeria Cave is included in the list of Areas or Sites of Community Importance for the Atlantic biogeographical region as SCI ES-1300017 (Commission Decision 2010/ 43/EU of 22 December 2009 adopting, pursuant to Council Directive 92/43/EEC).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Dyspnoi |
|
Family |
|
|
Genus |