
Indopinnixa haematosticta ( Sakai, 1934 ) Komai & Naruse & Yokooka & Taru & Shimetsugu & Watanabe, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5100.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:983DC53E-A4DA-4B94-AEBF-7A0678451A44 |
|
DOI |
https://doi.org/10.5281/zenodo.6314247 |
|
persistent identifier |
https://treatment.plazi.org/id/AC024E6B-FFF8-FFCE-FF29-5258463FFAE5 |
|
treatment provided by |
Plazi |
|
scientific name |
Indopinnixa haematosticta ( Sakai, 1934 ) |
| status |
comb. nov. |
Indopinnixa haematosticta ( Sakai, 1934) View in CoL n. comb.
( Figs. 2–8 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 , 16 View FIGURE 16 )
Pinnixa haematosticta Sakai, 1934: 42 View in CoL , fig. 3 [type locality: off Kisami, Shimoda, Izu Peninsula]; 1935: 203, fig. 107, pl. 57, fig. 4 (holotype); 1939: 600, text-fig. 85, pl. LXX, fig. 4 (holotype); 1976: 585, fig. 323, pl. 202, fig. 3 (holotype).— Yamauchi & Konishi 2005: 1241, fig. 1F (holotype).— Komatsu & Takeda 2009: 201 (off Ogi, Noto-cho, Ishikawa Prefecture; off Taniyama , Kagoshima Prefecture), 203 (key) .
Pinnixa hematosticta . — Ng et al. 2008: 247 (list).
Not Pinnixa haematosticta: Miyake 1983: 155 View in CoL , pl. 52-4 (Ezura, Nanki-shirahama, Wakayama Prefecture). –– Watanabe 2012: 217, colour fig. (Matsushima, Amakusa, Kumamoto Prefecture).
Records need to be verified:
Pinnixa haematosticta View in CoL .— Miyake 1961: 175 (Ariake Sea).— Miyake et al. 1962: 130 (Ariake Sea).— Nagai, 1990: 116 (Nankishirahama, Wakayama Prefecture).— Marumura & Kosaka 2003: 68 (Tsuchiya, Ehime Prefecture).
Type material. Holotype: KPM-NH107899, female (2.9× 6.9 mm), off Kisami , Shimoda, Izu Peninsula, Japan, 20 fathoms, sandy bottom, mixed with dead shells.
Other material examined. Honshu: CBM-ZC 15286, 1 male (2.5× 5.3 mm), Ena Bay, Miura Peninsula , Kanagawa Prefecture , intertidal, inhabiting tube of terebellid worm, 13 April 2013, digging with shovel, coll. T. Unagami and M. Taru; CBM-ZC 15287, 1 ovigerous female (4.2× 9.2 mm), same locality, intertidal, 26 July 2014, digging with shovel, coll. K. Yokoyama ; CBM-ZC 15288, 1 female (3.2× 7.4 mm), same locality, 7 June 2015, digging with trowel, coll. M. Taru ; RUMF-ZC-5948, 1 male (3.2× 6.5 mm), Tagoshi River estuary, Zushi , Kanagawa Prefecture, 23 April 2019, coll. H. Nakajima ; RUMF-ZC-5949, 1 male (2.7× 5.8 mm), Aburatsubo, Miura , Kanagawa Prefecture, 24 May 2019, coll. H. Nakajima ; CBM-ZC 15290, 2 females (3.5×7.6, 3.5× 7.8 mm, latter specimen DNA voucher), Tsumeki-zaki, Shimoda , Shizuoka Prefecture, shallow subtidal, 21 September 2016, yabby pump, coll. H. Yokooka ; CBM-ZC 15291, 1 male (3.6× 8.3 mm), 1 female (4.1× 8.5 mm), same locality, 3 December 2016, yabby pump, coll. H. Yokooka ; CBM-ZC 15289, 1 androgynous specimen (2.6× 5.6 mm), Kujyuppama, Shimoda , Shizuoka Prefecture, sand flat, 16 September 2016, coll. H. Yokooka ; CBM-ZC 15292, 1 androgynous specimen (2.7× 5.6 mm), Uramura, Toba , Mie Prefecture, intertidal, 20 June 2015, digging, coll. M. Shimetsugu ; CBM-ZC 15293, 1 female (2.7× 5.8 mm), same locality, 2 April 2017, digging, coll. M. Shimetsugu ; CBM-ZC 9135, 1 androgynous specimen (2.5× 4.7 mm), Fukaura, Kasado Island, Shimomatsu , Yamaguchi Prefecture, intertidal sand flat, 13 October 2002, digging with shovel, coll. H. Fukuda. Kyushu : RUMF-ZC-6000, 3 ovigerous females (2.8×5.8, 3.0×6.1, 3.1× 6.8 mm, second specimen DNA voucher), Kamenokojima, Sasebo , Nagasaki Prefecture, 33.1259N, 129.6783E, 3 October 2017, coll. T. Naruse & P. K. L. Ng GoogleMaps ; CBM-ZC 15294, 2 females (2.8×6.0, 2.8× 6.4 mm), Aitsu, Matsushima, Kami-Amakusa , Kumamoto Prefecture, intertidal, 21 April 2004, coll. T. Watanabe ; CBM-ZC 15295, 1 ovigerous female (2.8× 6.6 mm), same locality, 11 July 2006, coll. T. Watanabe ; CBM-ZC 15296, 1 female (2.3× 5.7 mm), same locality, 12 March 2008, coll. T. Watanabe ; NSMT-Cr 6470, 2 males (3.0×6.4, 3.5× 7.4 mm), 3 females (3.3×7.0–3.6× 7.5 mm), off Taniyama, Kagoshima Bay , 30–50 m, 7 May 1970 ; RUMF-ZC-5955, 9 males (1.8×3.3–2.9× 6.1 mm), 5 females (1.9×3.7–3.1× 6.4 mm), 15 ovigerous females (1.9×3.9–3.0× 6.8 mm), Fukiage Beach, Kominato, Minami Satsuma , Satsuma Peninsula , Kagoshima Prefecture, beach combing after typhoon, 20 September 2016, coll. D. Uyeno ; CBM-ZC 16586, 3 males (2.7×5.5–2.9× 6.1 mm), 3 ovigerous females (2.9×6.0, 3.2×6.6, 3.3× 6.9 mm, first specimen DNA voucher), same data ; CBM-ZC 15297, 1 male (2.5× 5.5 mm), 1 female (4.0× 9.1 mm), Kumano Fishing Port, Naka-Tane, Tanegashima Island , 0.5 m at low tide, 18 May 2018, yabby pump, coll. T. Komai .
Redescription. Male. Body and appendages well-sclerotized.
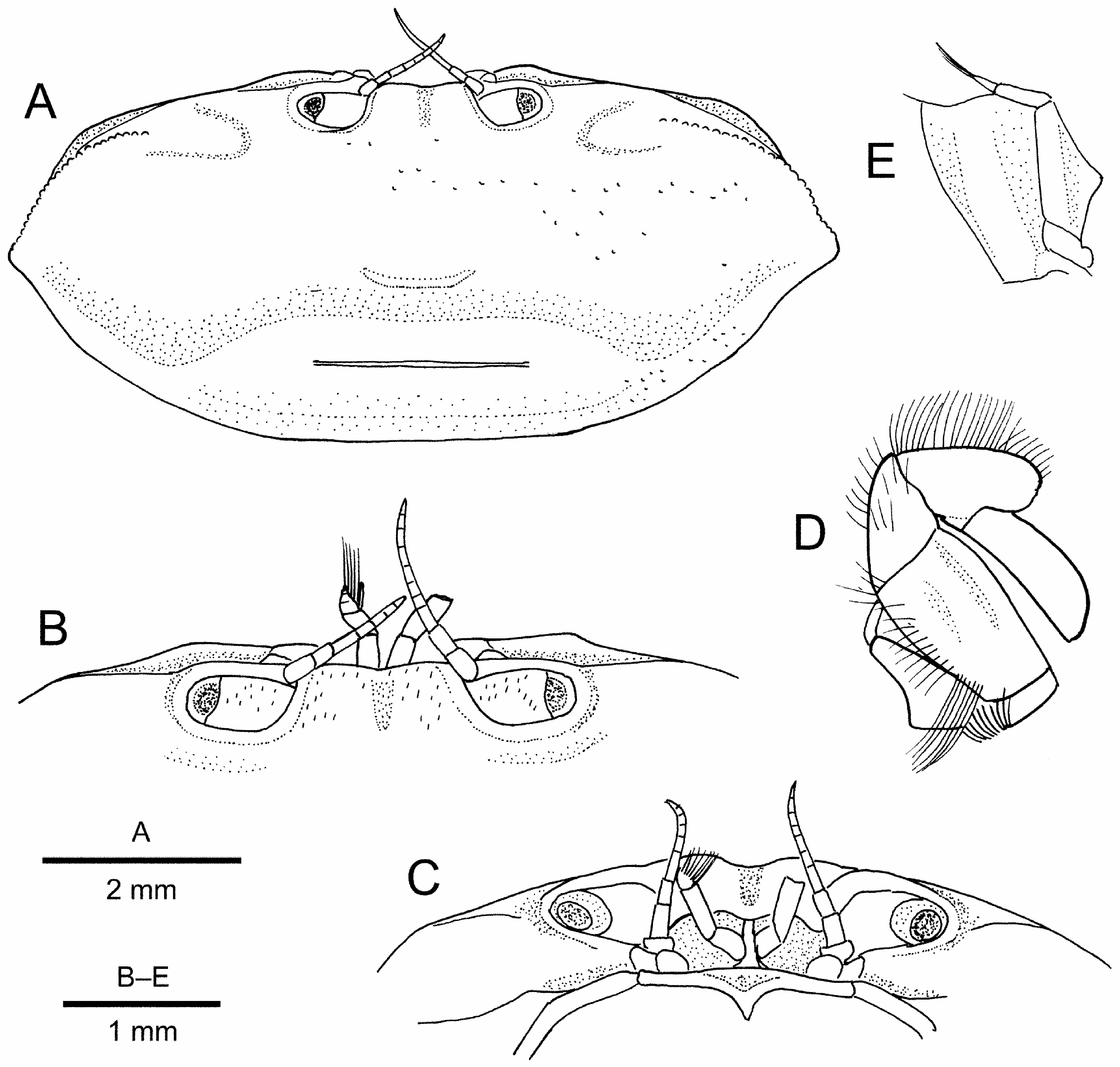
Carapace ( Figs. 2A View FIGURE 2 ; 3–C View FIGURE 3 ) transversely ovate, 2.1–2.3 times wider than long; dorsal surface gently convex in general, sparsely punctate, with very short, pigmented setae on front and adjacent to anterior margin; regions poorly defined, only shallow, median gastrocardiac groove discernible; cardiac region transversely elevated, with low but clearly delimited transverse ridge falling far short of posterolateral margins. Front slightly bilobed, slightly more than 0.1 times carapace width; lateral angle bluntly angular. Supraorbital margin entire, slightly limbed laterally, confluent with suborbital margin; suborbital margin smooth, entire. Hepatic region slightly swollen. Anterolateral margin gently arcuate, defined by granular ridge not confluent with supraorbital margin, starting some distance from orbit, forming distinct projection at juncture with posterolateral margin; posterolateral margin smooth, slightly sinuous, distinctly converging towards almost straight posterior carapace margin. Subhepatic region with low oblique ridge obscured by numerous short, plumose setae; suborbital region narrow; pterygostomial regions smooth.
Ocular peduncle ( Figs. 2A View FIGURE 2 , 3A–C View FIGURE 3 ) filling orbit, stout, with scattered very short, darkly pigmented setae on dorsal surface; cornea narrower than ocular peduncle, darkly pigmented.
Antennule ( Fig. 3B, C View FIGURE 3 ) with basal articles sub-globular, separated by median septum; distal 2 articles folded transversely; article 3 slightly longer than article 2; upper and lower flagella both shorter than article 3. Antenna with subquadrate basal article in contact with orbit; flagellum short.
Epistome ( Fig. 3C View FIGURE 3 ) longitudinally very narrow, posteromedian margin protruded into buccal space as short ridge; anterior buccal flame gently convex on either side of median ridge, with distinct suture at base of antennal article 1.
Maxillipeds 3 with large propodi and dactyli ( Fig. 3D, E View FIGURE 3 ), endopod and its setae filling most of buccal space. Ischium and merus completely fused to form subtriangular plate (= ischiomerus), external surface without median sulcus, but keeled along inner margin, outer margin obscured by numerous short to long plumose setae. Carpus shorter than ischiomerus, outer surface with numerous short plumose setae. Propodus subovate, spatuliform, with scattered short plumose setae on external surface. Dactylus also spatuliform, articulated at middle portion of flexor margin of propodus, widest at distal 0.25 length, distal part wider than proximal, tip rounded. Inner margins of ischiomerus and carpus, inner to distal margin of propodus, and inner margin of dactylus with very long, thick setae for filter feeding. Exopod completely hidden by outer margin of ischiomerus, shorter than ischiomerus, tapering distally, outer margin with subtriangular lobiform structure at about midlength; flagellum much shorter than exopod.
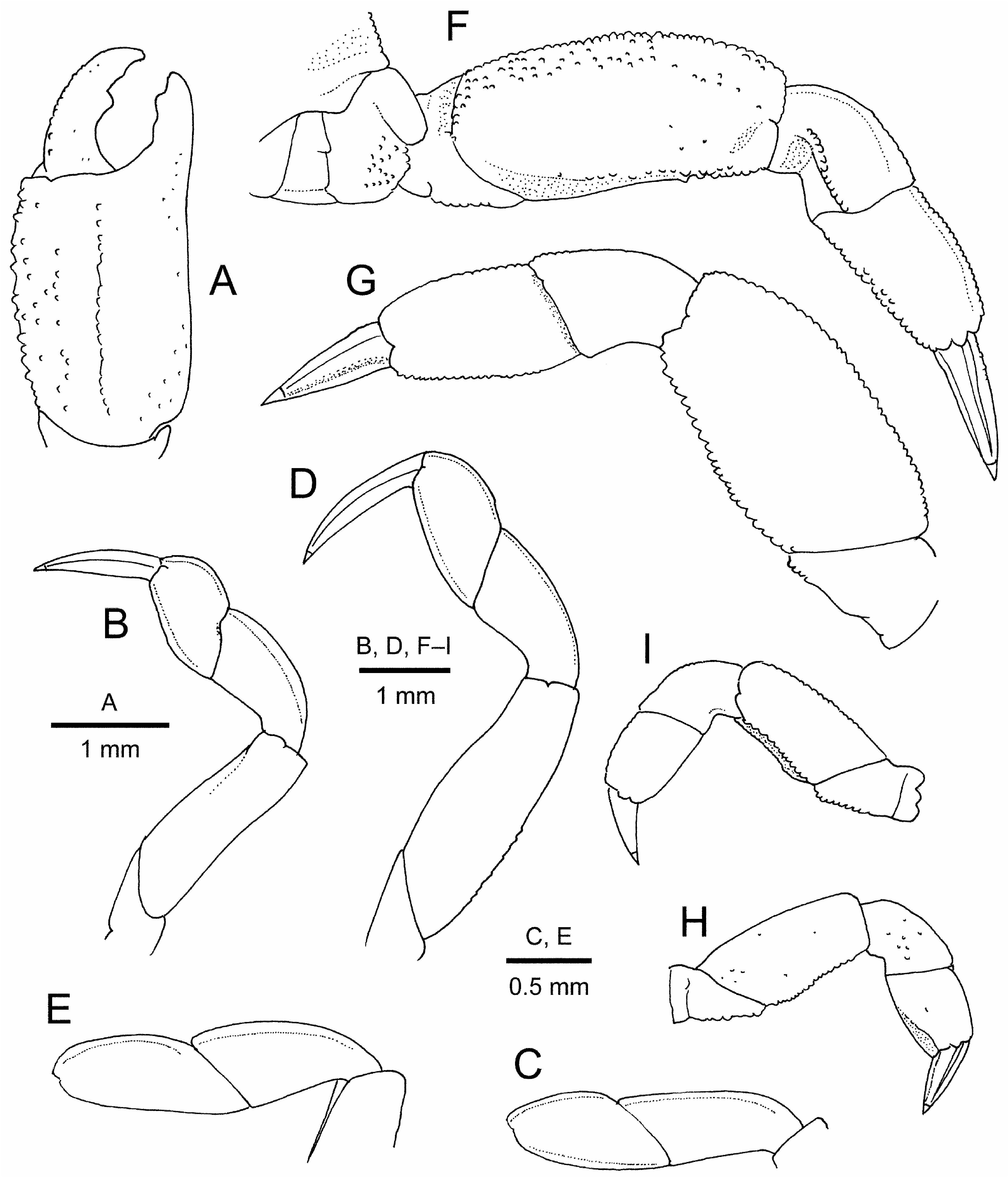
Chelipeds ( Figs. 2A, B View FIGURE 2 ; 4A View FIGURE 4 ) not enlarged; subequal and similar. Basis and ischium fused but suture visible, surfaces almost glabrous. Merus relatively short, without conspicuous armature, most surfaces obscured by plumose setae; upper surface almost flat, with few granules on proximal outer portion; inner margin convex, with row of granules; outer surface rounded; lower surface glabrous, with row of granules along outer side. Carpus short, cupshaped, also without conspicuous armature; upper surface with few short setae. Chela 1.9–2.2 times as long as high, fairly compressed; lower margin slightly sinuous. Palm 1.3–1.4 times as long as high; upper margin delimited by row of granules, obscured by numerous setae; outer surface with numerous short plumose setae obscuring scattered granules on upper half and having median longitudinal row of granules accompanied by short setae; inner surface with sparse, very short setae on dorsal side and numerous short to long plumose setae adjacent to lower margin; lower margin rounded, not carinate; fingers with wide hiatus filled by plumose setae; fixed finger nearly straight, not deflexed, terminating in small curved tip, occlusal margin with 2 or 3 blunt teeth at least in distal half. Dactylus 0.8 times as long as palm, noticeably curved, leaving distinct hiatus when closed; upper margin smooth or with row of minute granules and numerous plumose setae, occlusal margin armed with 1 small tooth at midlength.
Ambulatory legs moderately long, pereopod 4 largest, pereopod 5 smallest. Pereopod 2 ( Fig. 2A, B View FIGURE 2 ; 4B, C View FIGURE 4 ) moderately slender. Basis-ischium fused article short, with few granules on posterior margin. Merus slightly recurved, slightly narrowed distally, subtriangular in cross section; anterior surface non-carinate, with row of plumose setae; posterior surface with row of setae on upper side, otherwise almost glabrous. Carpus with extensor margin sharply carinate, flexor margin rounded with row of plumose seta on upper side. Propodus almost glabrous, 0.9–1.4 times as long as wide, 0.5–0.7 length of carpus; extensor margin sharply carinate, noticeably convex in large specimens; flexor margin also sharply carinate, less convex than upper margin. Dactylus nearly straight, 1.5 times as long as propodus, tapering to small corneous tip, with 4 distinct longitudinal carinae (extensor and flexor margins sharply carinate, upper and lower surfaces also each with median carina); extensor margin with short sparse setae.
Pereopod 3 ( Figs. 2A, B View FIGURE 2 ; 4D, E View FIGURE 4 ) moderately slender. Basis-ischium fused article short, with some granules on posterior face. Merus compressed, anterior margin bluntly carinate, nearly straight, with row of short to long plumose setae over entire length and row of small granules at least on proximal half; posterior surface bicarinate distally, with row of plumose setae on over entire length and row of granules at least on proximal half of upper side; upper and lower surfaces glabrous. Carpus with extensor margin gently convex and sharply carinate; flexor surface rounded with row of short to long plumose setae. Propodus 1.2–1.8 times as long as wide, almost glabrous except for row of short setae on flexor margin; extensor margin noticeably convex, sharply carinate; flexor margin gently convex, sharply carinate. Dactylus nearly straight, 1.5 times as long as propodus, tapering to small corneous tip, with 4 distinct longitudinal carinae (extensor and flexor margins sharply carinate, outer and inner surfaces also each with median carina); upper surface with row of short stiff setae on space between extensor and upper carinae.
Pereopod 4 ( Figs. 2A, B View FIGURE 2 ; 4F, G View FIGURE 4 ) stout, large, fairly compressed. Coxa enlarged, with prominent, glabrous lobe anteroventrally. Basis-ischium fused article with flat posterior surface, with few granules distally. Merus subovate, compressed, 1.7–2.0 times longer than wide; anterior margin gently convex, carinate, with rows of coarse granules and of plumose setae; upper surface with scattered coarse granules adjacent to anterior margin, obscured by plumose setae; lower surface smooth, glabrous; posterior surface concealed by dense plumose setae, flanked by carinae, upper carina delimited with single or double row of granules, becoming obsolete proximally, and with row of plumose setae; lower carina distinct over entire length, slightly sinuous, bordered with coarse granules. Carpus short, widened distally; extensor margin sharply carinate; upper surface with scattered short plumose setae; lower surface glabrous; flexor surface forming shallow concavity accommodating posterodistal angle of merus, defined on upper side by row of granules. Propodus tapering distally, 1.6 times as long as wide; extensor margin slightly convex, sharply carinate; upper surface with scattered short setae; lower surface glabrous; flexor surface flanked by sharp, granulate carinae, with dense plumose setae. Dactylus almost straight, subequal in length to propodus, tapering to small corneous claw, with 5 longitudinal carinae and sparse short setae; flexor surface narrow, with 2 longitudinal rows of short setae.
Pereopod 5 ( Figs. 2A, B View FIGURE 2 ; 4H View FIGURE 4 ) short, fairly compressed, falling far short of distal margin of merus of pereopod 4 when extended, with numerous long plumose setae on margins. Coxa with prominent knob-like protuberance on dorsodistal margin. Basis-ischium fused article with few granules on posterior surface. Merus twice as long as wide, anterior margin bluntly carinate, granulate; upper and lower surfaces almost glabrous; posterior surface flanked by sharp, granulate carinae, with numerous plumose setae. Carpus extensor margin non-carinate; upper and lower surfaces almost glabrous. Propodus subquadrate, 1.2 times as long as wide, extensor margin not carinate; upper and lower surfaces almost glabrous; posterior surface thickly setose. Dactylus subequal in length to propodus, subconical, with 2 rows of short stiff setae on extensor margin.
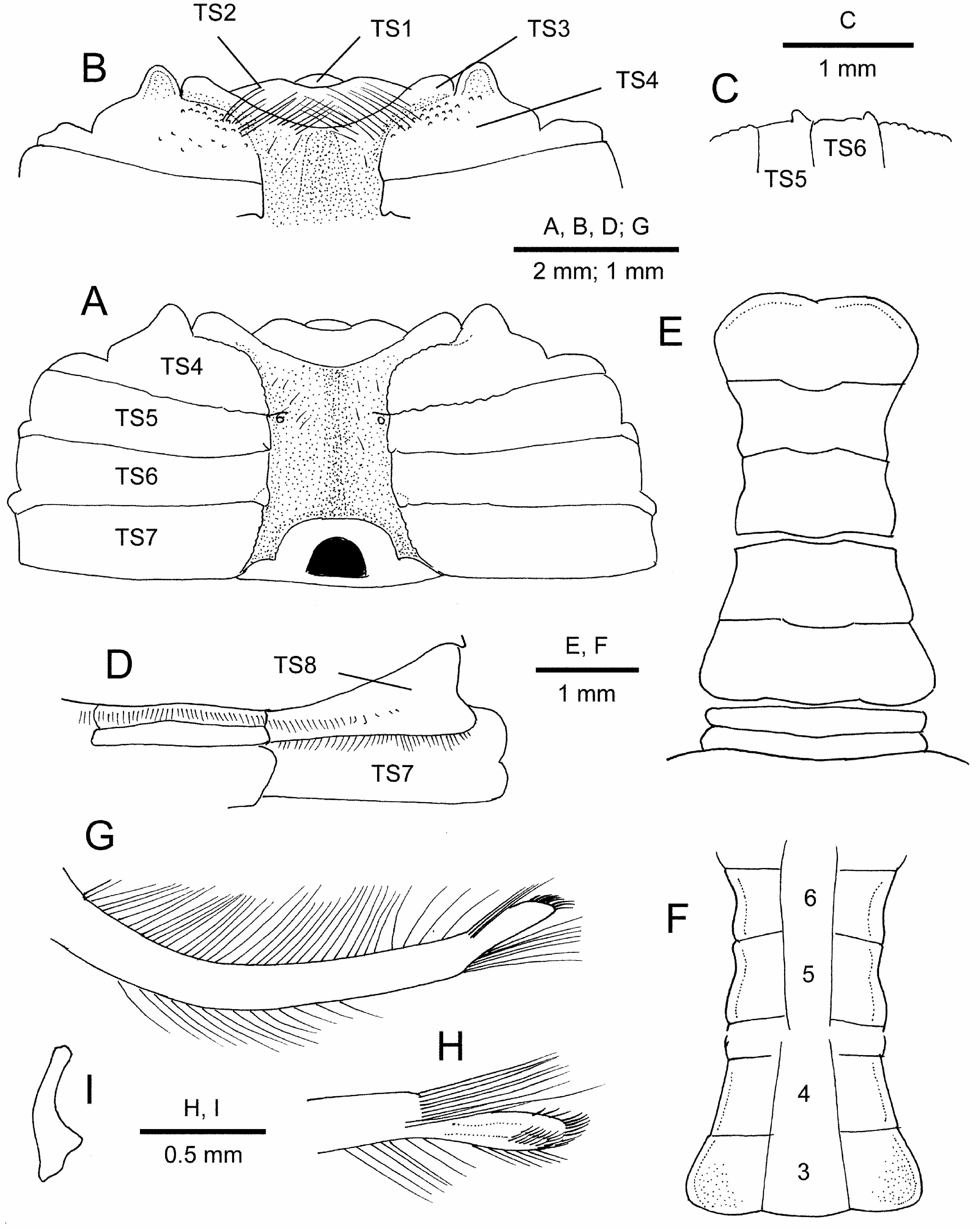
Thoracic sternum ( Figs. 2B View FIGURE 2 ; 5A, B, D View FIGURE 5 ) wide, polished, edges of sternites abutting sutures granular or smooth. Sternites 1 and 2 incompletely fused, deeply sunken into buccal cavity together with anteromedian part of sternite 3, obscured by thick setae; only lateral parts of sternite 3 clearly visible. Sternite 3 and 4 clearly separated by distinct, granulate ridge; distal part of gonopod 1 fitted into oblique lateral part of sternite 3. Sternite 4 with granules anteriorly; each anterolateral margin produced into rounded lobe, separated by anterolateral angle of sternite 3 by deep notch. Sternites 5–7 almost smooth, sternite 7 longitudinally widest. Sternite 8 wide, fully exposed, widened toward lateral margin, nearly perpendicular against horizontal plane of carapace, but partially visible in dorsal view, with transverse rows of setae, not extending to lateral margin, on midline and on morphologically anterior margin along suture, extending over entire width. Sterno-pleonal cavity deep, reaching to base of buccal cavity, marked by distinct ridges where sternites fit against lateral margins of closed pleon; margins of sterno-pleonal cavity along sternites 4–6 smooth, not granular; those on sternites 5 and 6 posteriorly raised to form prominent tubercles that grip closed pleon ( Fig. 5C View FIGURE 5 ); press-button pleonal locking mechanism reduced into tiny tubercle inside of sterno-pleonal cavity of sternite 5 near suture between sternites 4/5. Penis sternal.
Pleon ( Figs. 2B View FIGURE 2 ; 5E, F View FIGURE 5 ) narrow, constricted, outer surface polished. Somites 1 and 2 transversely wide and very short, somite 1 with transverse row of short setae on midline. Somite 3 widest, trapezoidal, lateral margins rounded proximally. Somites 4–6 functionally fused, immobile although sutures clearly visible; somite 4 trapezoidal, lateral margins with shallow concavity corresponding to ledge-like structure on thoracic sternite 6; somite 5 subquadrate, lateral margins sinuous, concavities corresponding to ledge-like structure on thoracic sternite 5; somite 6 widened distally, with lateral margins concave; gonopodal plate absent. Telson covering posterior part of buccal cavity, almost as wide as pleomere 3, about 2 times as wide as long; lateral margin strongly convex, distal margin bilobed with shallow median notch.
Male gonopod 1 ( Fig. 5G, H View FIGURE 5 ) relatively long, somewhat compressed, reaching posterior end of buccal cavity, gently curved outward, outer and inner margins with row of numerous long, plumose setae; distal process delimited by slight constriction at base, strongly compressed, faintly recurved, terminal margin blunt, with patch of dense very short stiff setae on distal half of dorsal surface; shoulder at base of distal process with tuft of numerous long setae reaching to terminus of distal process. Male gonopod 2 very short, gently curved, with slightly inflated, spatuliform tip, less than 0.2 length of gonopod 1; base strongly inflated.
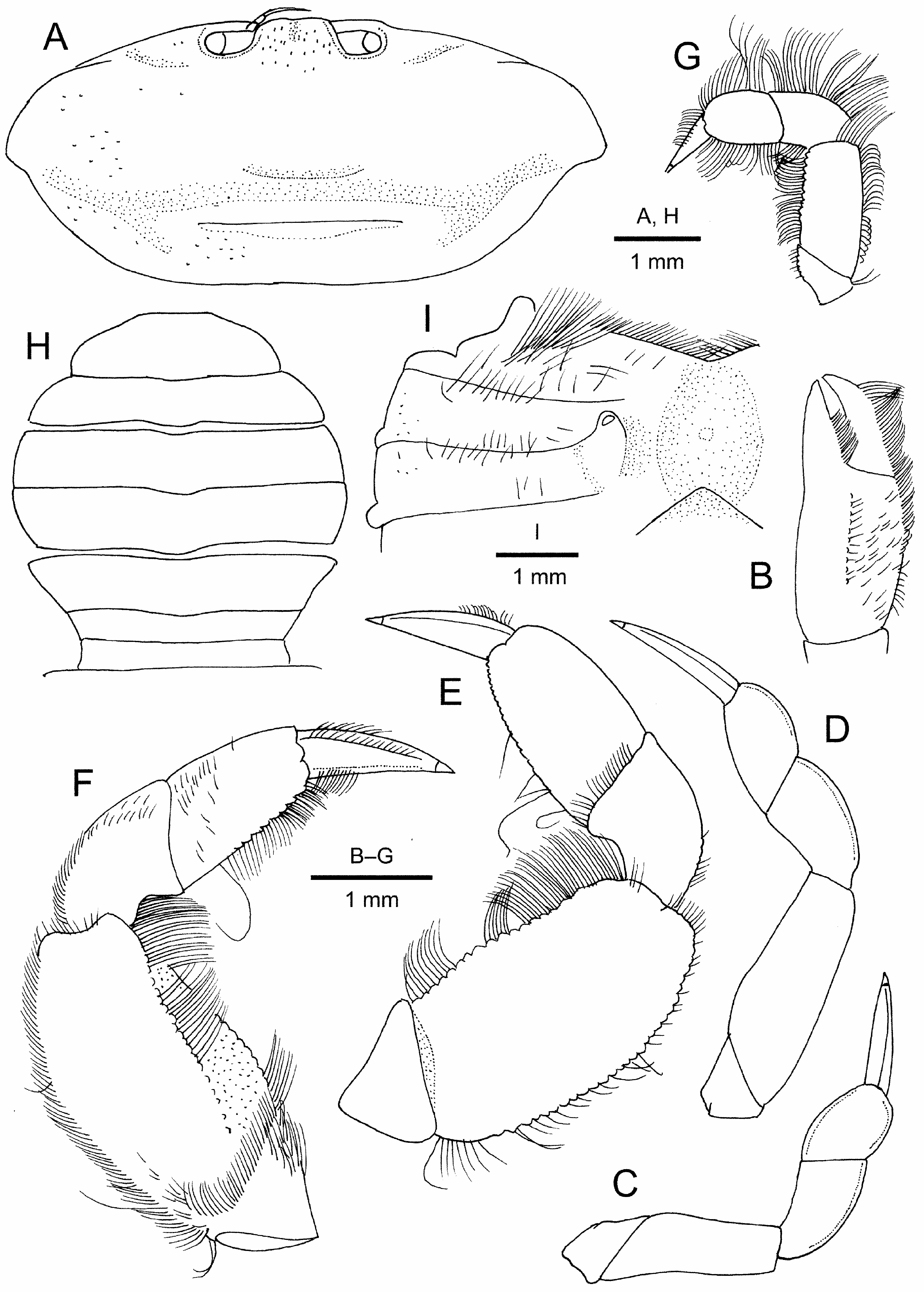
Female. Carapace ( Figs. 6A View FIGURE 6 ; 7A View FIGURE 7 ) generally similar to that of males (2.2 times as wide as long in holotype), but anterolateral margins slightly more sloping; cardiac region usually with distinct transverse ridge.
Chela ( Figs. 6A, B View FIGURE 6 ; 7B View FIGURE 7 ) proportionally smaller than in males, 2.3–2.5 times as long as high; lower margin nearly straight or faintly sinuous. Palm 1.4–1.6 times as long as high; upper margin with row of granules; outer surface with scattered granules superior to median line of setae; occlusal margin of fixed finger with row of 3–5 small blunt teeth. Dactylus 0.7–0.8 times as long as palm, curved distally, terminating in acute tip; upper margin gently arcuate, with row of granules; occlusal margin with 1 small but conspicuous tooth at about mid-length, and with row of minute teeth distal to median tooth.
Pereopods 2–5 relatively shorter than in males (cf. Fig. 6A, B View FIGURE 6 versus Fig. 2A, B View FIGURE 2 ; Fig. 7C–G View FIGURE 7 ). Pereopod 2 propodus slightly wider than long (1.1 times as wide as long in holotype) to 1.3 times as long as wide. Pereopod 3 propodus 1.3–1.6 times as long as wide. Pereopod 4 merus 1.7–1.9 times as long as wide.
Female thoracic sternum ( Fig. 6B View FIGURE 6 ) with most surfaces smooth or only slightly punctate, sometimes with sparse setae partially; lateral parts of sternites 4–8 exposed, not concealed by closed pleon (but developed setae on lateral margins of pleon sometimes covering lateral parts of sternite). Sternites 3 and 4 completely fused; widely concave anterior margin fringed with dense setae. Gonopore tubercle (vulva) small, low, generally rounded, extending anteriorly across half-length of sternite 6, overreaching more than half length of sternite 5; gonopore rounded ( Fig. 7I View FIGURE 7 ).
Pleon ( Figs. 6B View FIGURE 6 ; 7H View FIGURE 7 ) relatively narrow (lateral parts of thoracic sternum exposed), subcircular, somite 4 widest; in ovigerous females, lateral margins fringed with thick setae covering lateral parts of thoracic sternum. Telson ( Fig. 7H View FIGURE 7 ) 3.1–3.3 times as wide as long, reaching to bases of maxilliped 3, distal margin straight or slightly concave.
Colouration in life. Base color of body and appendages gray-brown or brown, sometimes mottled, on dorsal side, whitish or pale yellow-brown on ventral side; carapace dorsal surface occasionally with reddish brown patches; thoracic sternites, pleon and lower surfaces of pereopods occasionally with scattered brown or gray spots; setae on pereopods generally brown ( Figs. 6A, B View FIGURE 6 ; 8A, B View FIGURE 8 ).
Size. Males 1.8×3.3–3.6× 8.3 mm; ovigerous females 1.9×3.9–4.2× 9.2 mm.
Abnormalities. Three specimens [CBM-ZC 9135 (2.5× 4.7 mm), 15289 (2.6× 5.6 mm) and 15292 (2.7× 5.6 mm)] are androgynous in having male-like pleons (cf. Fig. 8B View FIGURE 8 ) with functionally fused pleomeres 4–6, which are proportionally slightly wider than in normal males, gonopores on the thoracic sternite 6, and pleomeres 2–5, of which the anterior two pairs are biramous and posterior two pairs are uniramous; the sterno-pleonal cavity is proportionally wider than in normal males, defined by less developed ridges; rudimentary press buttons are evident. The size of these androgynous specimens are included in the range of the adult size of the species (see above “Size”). Androgyny in the Brachyura is little known, but some examples have been reported in the pinnotherid Pinnotherinae De Haan, 1833 (e.g., Manning 1993, 1998; Ahyong and Ng 2008; Ahyong 2020), the varunid Pseudopinnixa Ortmann, 1894 ( Komai & Konishi 2012), and the ocypodid Leptuca Bott, 1973 ( Zou & Fingerman 2000), the potamid Geothelphusa Stimpson, 1858 (e.g. Takahashi et al. 2000), and portunid Catoptrus A. Milne-Edwards, 1870 ( Naruse & Uyeno 2021). In the case of Indopinnixa haematosticta n. comb., there is no doubt that the observed androgyny represents an abnormality.
Distribution and habitat. So far known only from Japan: Kanagawa Prefecture to Tanegashima Island in Ohsumi Islands, southern Kyushu (Pacific side) and Yamaguchi Prefecture (Sea of Japan side); intertidal to 50 m.
Most specimens examined were extracted from sand substrates by using yabby pumps or shovels (see “Material examined”). One male specimen (CBM-ZC 15286; Ena Bay, Miura, Kanagawa Prefecture) was collected from a tube of terebellid worm, representing an example of possible association.
Remarks. The female holotype of Indopinnixa haematosticta n. comb. is now in rather poor condition: the entire cephalothorax, pleon and appendages are decalcified; the right cheliped is missing; setae on appendages thickly bear mucus-like matter, sometimes making difficult to observe detailed structure of those appendages; in particular, the maxillipeds 3 are completely obscured, which is difficult to remove without much damage to the specimen. Fortunately, Sakai (1934, 1939, 1976) provided a figure of the maxilliped 3. The following features are still visible in the holotype: carapace broad (> 2.0 times as long as broad), with sharply delimited transverse carina, falling far short of lateral margins of carapace, on weakly elevated cardiac region ( Fig. 7A View FIGURE 7 ); chela with lines of numerous setae on upper and lower margins and along mid-line of outer surface of palm ( Fig. 7C View FIGURE 7 ); fixed finger of chela not deflexed, straight ( Fig. 7C View FIGURE 7 ); cheliped dactylus nearly straight except for curved tip, shorter than palm ( Fig. 7C View FIGURE 7 ); carpi of pereopods 2 and 3 each with sharply carinate extensor margin ( Fig. 7D, E View FIGURE 7 ); propodi of pereopods 2 and 3 each with sharply carinate extensor and flexor margins ( Fig. 7D, E View FIGURE 7 ); merus of pereopod 4 stout, 1.8 times as long as wide, posterior surface filled with dense plumose setae ( Fig. 7F, G View FIGURE 7 ); pleomere 1 with fringe of short setae extending onto thoracic sternite 8 as transverse row (setae omitted in Fig. 7E View FIGURE 7 , but see Fig. 6B View FIGURE 6 for supplemental information). We identified the newly collected material from Japanese mainland used in this study based on these features. Yamauchi & Konishi (2005) failed to detect a sharply delimited transverse carina on the cardiac region of the carapace in the holotype, but we confirmed its presence, as specifically mentioned and illustrated by Sakai (1934, 1935, 1939, 1976). Examination of the male specimens has clarified that this species has functionally fused pleomeres 4–6, and this led the authors to assign Pinnixa haematosticta to Indopinnixa . Additional characters that are potentially generically diagnostic, shared with the other species of Indopinnixa include: the maxilliped 3 dactylus is large and heavy, longer than the propodus ( Fig. 3D, E View FIGURE 3 ); the external surface of the chela palm bears longitudinal ridges or lines of granules on the midline ( Fig. 5A View FIGURE 5 ); the pereopod 4 is very stout ( Figs. 4F, G View FIGURE 4 ; 7F, G View FIGURE 7 ); the male pleon is devoid of a gonopodal plate developed on the inner surface ( Fig. 5F View FIGURE 5 ); the telson does not taper, being wider than the narrowest portion of the pleomere 6 ( Fig. 5E View FIGURE 5 ).
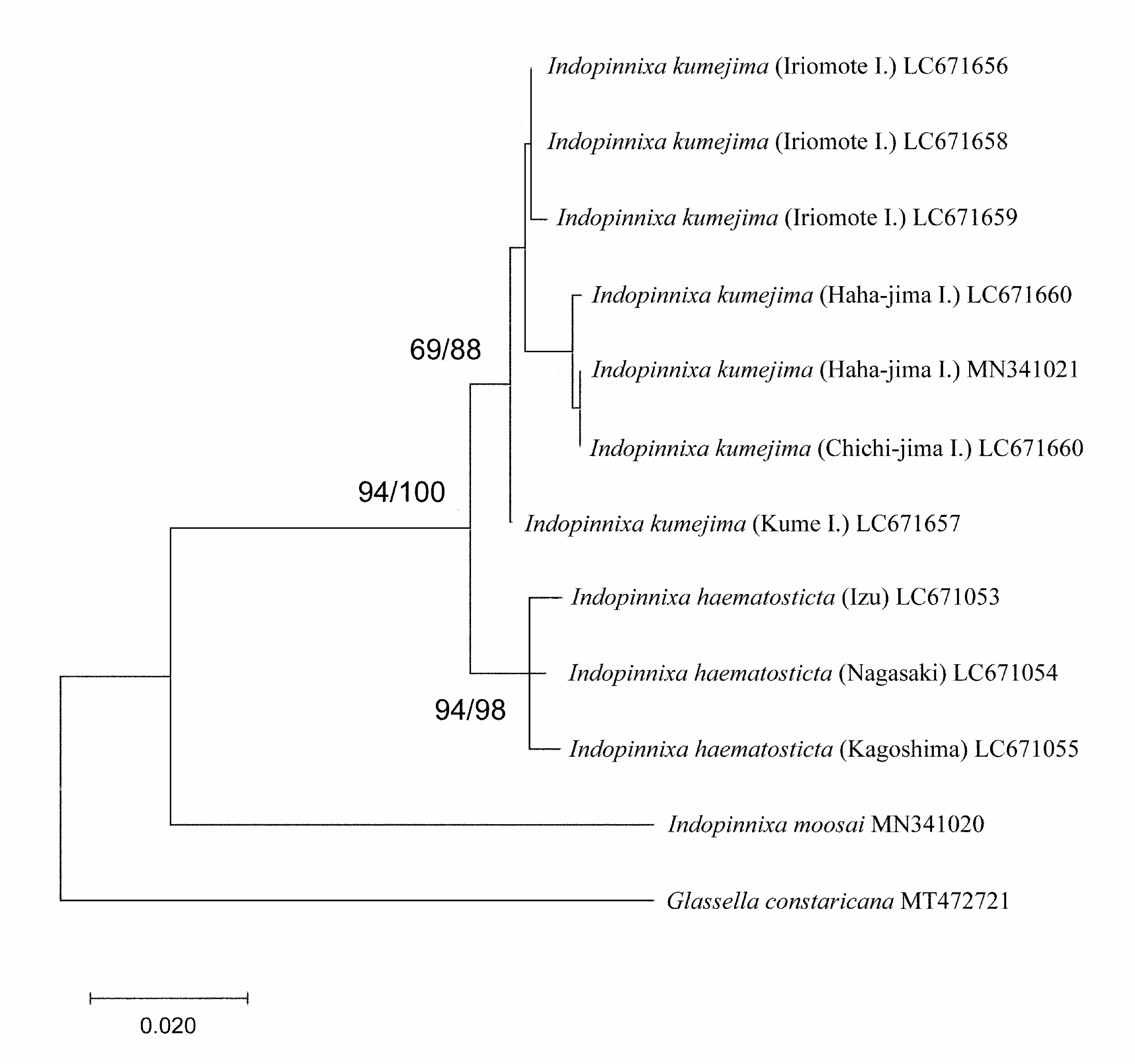
The assignment of the present species to Indopinnixa requires careful comparison with congeners, because in previous works describing species of Indopinnixa , I. haematosticta n. comb. was not considered for comparison ( Manning & Morton 1987; Davie 1992; Rahayu & Ng 2010; Naruse & Maenosono 2012; Ng 2014). The proportionally wider carapace (2.0–2.3 times as wide as long) usually with a distinct transverse carina on the cardiac region distinguishes I. haematosticta n. comb. from all but I. kumejima . In I. kasijani , I. moosai , I. mortoni , I. oryza and I. sipunculana , the carapace is twice or less than twice as long as wide, even in adults; there is no conspicuous transverse carina on the cardiac region of the carapace (cf. Manning & Morton 1987; Davie 1992; Rahayu & Ng 2010; Naruse & Maenosono 2012). Indopinnixa shellorum has a distinct transverse carina on the cardiac region of the carapace, but the carapace is distinctly proportionally narrower (1.6 times as wide as long; cf. Ng 2014) than in I. haematosticta n. comb., and the eyes are relatively larger in I. shellorum than in I. haematosticta n. comb. (cf. Ng 2014, fig. 3A versus Figs. 3A View FIGURE 3 ; 7A View FIGURE 7 ). The shape of the distal process of the male first gonopod is also different between I. haematosticta and the other five species: in I. mortoni (cf. Davie 1992: fig. 1D, E), I. oryza (cf. Naruse & Maenosono 2012: fig. 6b, c) and I. sipunculana (cf. Manning & Morton 1987: fig. 1F, G), the distal process is tube-like, almost straight; in I. kasijani (cf. Rahayu & Ng 2010: fig. 3A, B) and I. shellorum (cf. Ng 2014: fig. 3I, J, K), the distal process is also tube-like, but recurved; in I. moosai (cf. Rahayu & Ng 2010: fig. 6A–D), the distal process is thick with a rounded terminal margin.
Indopinnixa kumejima is morphologically very similar to I. haematosticta n. comb., sharing all diagnostic characters mentioned above. Differentiating characters between the two species are discussed under the account of I. kumejima .
During this study, specimens identified with P. haematosticta n. comb. by Komatsu & Takeda (2009) have been reexamined, and the specific identify was confirmed.
Miyake (1983) identified one female specimen from Nanki-Shirahama, Wakayama Prefecture, with Pinnixa haematosticta . However, according to the given picture ( Miyake 1983: pl. 52, fig. 4), the pereopod 4 merus is devoid of a thick field of setae on the posterior surface, which is characteristic of I. haematosticta n. comb.; and the carapace is 1.6 times as long as wide, being fairly proportionally narrower than in I. haematosticta n. comb. rediagnosed herein. Consequently, it is considered that Miyake’s (1983) specimen represents a species other than I. haematosticta n. comb.
Identifications of the following records remain to be verified, because no voucher specimens have been available for examination or no information on the morphology was provided: Miyake (1961: Ariake Sea); Miyake et al. (1962: Ariake Sea); Nagai (1990: Nanki-shirahama, Wakayama Prefecture); Marumura & Kosaka (2003: Tsuchiya, Ehime Prefecture).
The identity of Pinnixa penultipedalis Stimpson, 1858 , must now be considered in view of the present understanding of I. haematosticta n. comb. The species was originally described on the basis of a single female specimen from Hong Kong ( Stimpson 1858). Posthumously, Stimpson (1907) gave a more detailed description of the species, but without any illustration. Records under this species name include Ortmann (1894; Nagasaki, Japan), Shen (1937; China), Barnard (1955; South Africa), Miyake (1961; Sea of Ariake, Japan), Miyake et al. (1962; Amakusa and Sea of Ariake, Japan), Watanabe & Tanida (2001; Harima-nada, Seto Inland Sea); Yamauchi & Konishi (2005; off Oki Island, Sea of Japan), and Youn et al. (2019; Korea). Watanabe (2012) changed the identification of Watanabe & Tanida (2001) to Pinnixa aff. penultipedalis because of the uncertainty of the identification. Some other workers have also mentioned on Pinnixa penultipedalis . For example, Sakai (1934; 1939) cited the accounts by the original author and the record by Ortmann (1894), and Sakai (1976) questioned the identification by Ortmann (1894). Dai et al. (1986) and Dai & Yang (1991) gave an account of the species, based on the account by Shen (1937). In the original description of Indopinnixa sipunculana, Manning & Morton (1987) compared their new taxon with P. penultipedalis . Naruse & Maenosono (2012) commented on the taxonomy of P. penultipedalis , suggesting that none of the subsequent records since Stimpson (1858, 1907) represent P. penultipedalis . The holotype of P. penultipedalis is no longer extant, because it was destroyed in the Chicago Fire of 1871 ( Evans 1967). It is not easy to assess the generic position of Stimpson’s (1858) taxon because of the lack of sufficient morphological details in the existing descriptions ( Stimpson 1858, 1907). Nevertheless, comparison with the description by Stimpson (1907) suggests that P. penultipedalis is not conspecific with I. haematosticta n. comb. because of the carapace having a transverse ridge near the posterior extremity traversing the whole breadth of the carapace and the pereopod 4 merus being fourfifths (= 0.8 times) as broad as long. In I. haematosticta n. comb., the carapace has a clearly delimited transverse ridge limited to the cardiac region; the merus of the pereopod 4 is about 0.4–0.6 times as broad as long. During this study, the male specimen identified with P. penultipedalis by Yamauchi & Konishi (2005; KPM 0000080) was reexamined. In conclusion, the specimen represents a different species from those referred to as P. penultipedalis by Shen (1937), Watanabe & Tanida (2001) and Youn et al. (2019) and possibly representing an undescribed species of Indopinnixa . This issue will be dealt with in a separate article. In order to fully establish the identity of P. penultipedalis , examination of material from the type locality, Hong Kong, which agrees with the description by Stimpson (1907), would be absolutely necessary. Recently, Wong et al. (2021) reported on a large amount of brachyuran material collected by trawling around Hong Kong, but P. penultipedalis was not rediscovered.



| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
Genus |
Indopinnixa haematosticta ( Sakai, 1934 )
| Komai, Tomoyuki, Naruse, Tohru, Yokooka, Hiroyuki, Taru, Masanori, Shimetsugu, Miho & Watanabe, Tetsuya 2022 |
Pinnixa hematosticta
| Ng, P. K. L. & Guinot, D. & Davie, P. J. F. 2008: 247 |
Pinnixa haematosticta: Miyake 1983: 155
| Watanabe, T. 2012: 217 |
| Miyake, S. 1983: 155 |
Pinnixa haematosticta
| Marumura, M. & Kosaka, A. 2003: 68 |
| Nagai, S. 1990: 116 |
| Miyake, S. & Sakai, K. & Nishikawa, S. 1962: 130 |
| Miyake, S. 1961: 175 |
Pinnixa haematosticta
| Komatsu, H. & Takeda, M. 2009: 201 |
| Yamauchi, T. & Konishi, K. 2005: 1241 |
| Sakai, T. 1934: 42 |