
Histionotophorus, EASTMAN, 1904
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00623.x |
|
DOI |
https://doi.org/10.5281/zenodo.10545587 |
|
persistent identifier |
https://treatment.plazi.org/id/2816A226-7754-9657-411A-1895FBC57E8A |
|
treatment provided by |
Valdenar |
|
scientific name |
Histionotophorus |
| status |
|
GENUS † HISTIONOTOPHORUS EASTMAN, 1904
† Histiocephalus De Zigno, 1887: 31 (type species Histiocephalus bassani De Zigno, 1887: 31 , by monotypy).
† Histionotophorus Eastman, 1904: 32 (replacement name for Histiocephalus De Zigno, 1887: 31 , preoccupied by Histiocephalus Diesing, 1851: 80 , 230, for a genus of nematodes, therefore taking the same type species, Histiocephalus bassani De Zigno, 1887: 31 ).
Diagnosis: A brachionichthyid genus unique and derived in having an elongate pointed caudal fin; foramina of the first dorsal-fin pterygiophore (which provide articulation for the illicium and second dorsal-fin spine) closely spaced; pterygiophores of the soft dorsal fin greatly enlarged; length of anal-fin base less than 20% SL; length of caudal peduncle greater than 15% SL. The genus is further distinguished from all other brachionichthyid genera in having the following combination of character states: jaw teeth small and appearing granular; vertebrae 19 (rarely 20); neural spines of abdominal vertebrae simple, not spatulate; haemal spines of abdominal vertebrae posteroventrally directed; first dorsal-fin pterygiophore greatly enlarged, hypertrophied; dorsal-fin rays 12–13; dorsal-fin rays extremely elongate; anal-fin rays eight to nine; pectoral-fin radials extremely elongate, length nearly 35% SL; body depth at origin of soft-dorsal fin nearly 42% SL; body depth at origin of anal fin approximately 31% SL; pectoral-fin rays seven; pelvic fin one spine and five rays; caudal-fin rays nine, the ventral-most ray greatly reduced; skin naked, without dermal spinules.
Included species: Monotypic.
† HISTIONOTOPHORUS BASSANI ( DE ZIGNO, 1887)
( FIGS 1–4 View Figure 1 View Figure 2 View Figure 3 View Figure 4 )
† Histiocephalus bassani De Zigno, 1887: 23–24 , pl. 1, fig. 9; Woodward, 1901: 579–580.
† Histionotophorus bassani Eastman, 1904: 32–33 , pl. 1, figs 1–3; Eastman, 1905: 30; Gill, 1905: 845–846; Regan, 1912: 283; Le Danois, 1964: 141–144, figs 75–76; Rosen & Patterson, 1969: 442, fig. 60, pl. 76, fig. 2; Pietsch, 1981: 416–417; Frickhinger, 1991: 695.
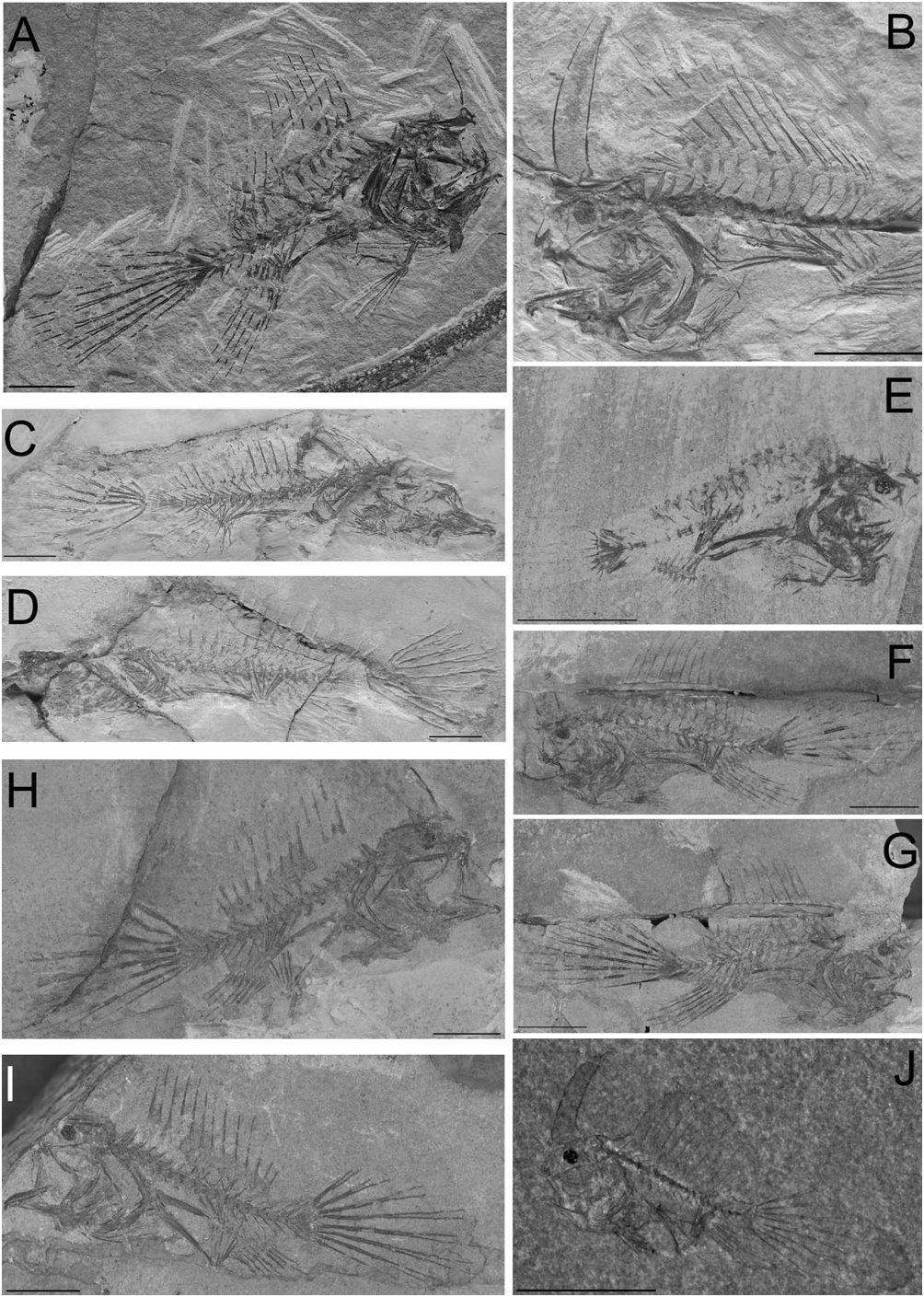
Holotype: MGPD 68487 , nearly complete skeleton in part and counterpart, 55.1 mm SL; late early Eocene , Ypresian ; Monte Bolca, Pesciara cave site.
Additional material: NHM 19060, complete skeleton, 16.4 mm SL, from the type locality; CM 5237 , incomplete skeleton lacking the anterior part of the head skeleton and distal portions of dorsal-fin spines and rays, caudal-fin rays preserved as impression only; from the type locality. MCSNV I.G.23163, complete skeleton, 50.5 mm SL, from the type locality; MCSNV S.30, incomplete skeleton lacking the head and anterior part of the axial skeleton, from the type locality; MCSNV T.162/163, incomplete and poorly preserved specimen, from the type locality; MCSNV T.358, incomplete specimen lacking the caudal portion of the skeleton, 38.5 mm estimated SL, from the type locality. MCZ 5176 About MCZ a/5176b, nearly complete skeleton in part and counterpart, 34.5 mm SL, from the type locality; MCZ 5177 About MCZ a/5177b, nearly complete skeleton in part and counterpart, 38.4 mm SL, from the type locality .
Diagnosis: As given for the genus.
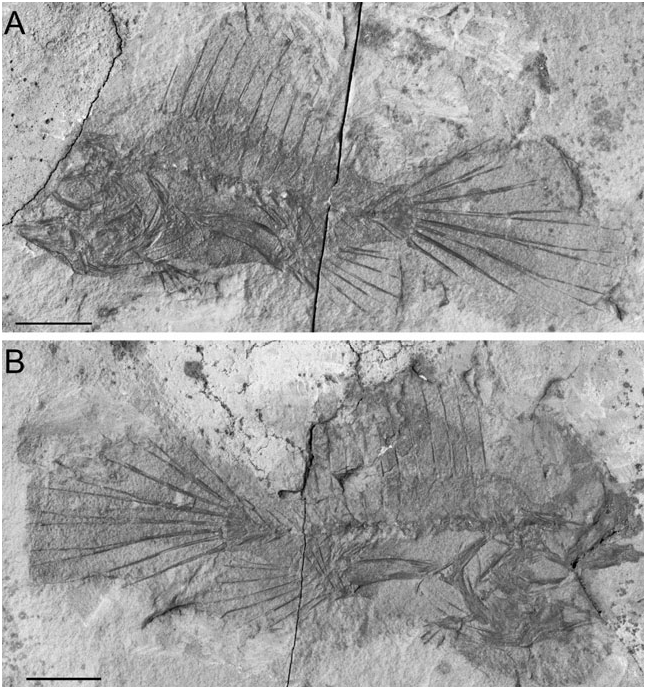
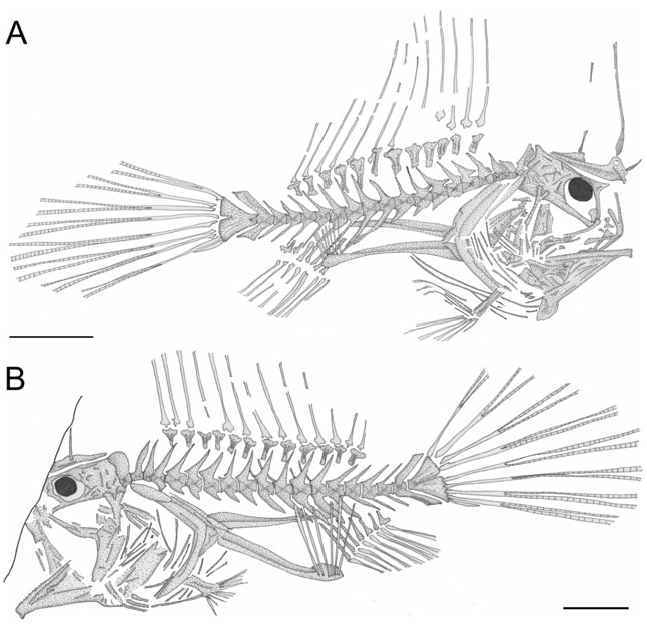
Description: The body is moderately elongate and slightly compressed ( Figs 1–3 View Figure 1 View Figure 2 View Figure 3 ). The dorsal and ventral profile of the anterior part of the body is convex. The abdomen is expanded ( Fig. 2B, F–G View Figure 2 ). The eyeball (based on preserved pigment) is close to the dorsal margin of the head. The caudal peduncle is well developed, its average length greater than 16% SL (see Table 1). The head is short, contained more than three times in standard length. The snout is short (10.1–14.6% SL). The eyeball is relatively small (5.9–8.5% SL). The mouth is nearly horizontal, terminal, and relatively small. The illicium is slender and terminal on the snout. The second and third dorsal-fin spines are interconnected by a membrane ( Fig. 2J View Figure 2 ). A membrane also connects the third spine with the dorsal mid-line of the body. The soft-dorsal fin is extremely well developed, sail-like. The central rays of the soft dorsal and anal fins are longer than the others; for this reason the posterior portion of the anal fin appears to be orientated vertically. The analfin base is relatively short (15.5% SL). The arm-like pectoral fins are characterized by extremely elongate radials (31.5–41.4% SL), bearing slender filamentous soft rays. The pelvic fin is elongate, with a short anterior spine and slender delicate soft rays. The caudal fin is sharply pointed ( Fig. 3A View Figure 3 ), elongate (as long as 63.7% SL). For additional counts and measurements, see Table 1.
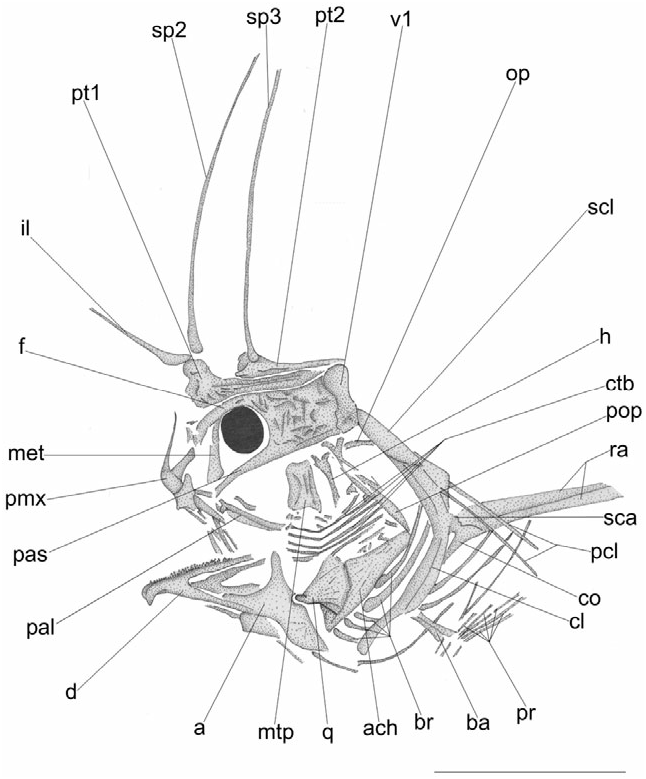
The neurocranium is relatively deep and robust ( Fig. 4 View Figure 4 ). The frontals are the largest bones of the skull roof, each relatively wide posteriorly, tapering anteriorly to form a prominent rostral process. The parietals are probably separated from each other. The mesethmoid, which forms a sort of interorbital septum, is columnar, with a broadly expanded ventral portion and a moderately constricted middle portion; the dorsal end of this bone articulates with the central extensions of the frontals. There is an evident gap between the mesethmoid and the vomer and parasphenoid that was probably occupied originally by the ethmoid cartilage. The lateral ethmoid is poorly preserved in all specimens examined; this bone appears to be thin and delicate, of irregular shape. The vomer is toothless, with a flattened ventral surface. The parasphenoid is a stout shaft throughout most of its length. The sphenotic (preserved in part only in MCSNV I.G. 23163) forms a prominent flange that extends laterally. The limits of the pterotic, prootic, epioccipital, and basioccipital are difficult to recognize as a result of inadequate preservation.
The premaxilla bears a long slender ascending process, a laminar well-developed articular process, and a thin spatulate postmaxillary process; the ascending and articular processes form an obtuse angle at their junctions. Minute granular teeth arranged in many irregular rows can be observed along the ventral surface of the alveolar process ( Figs 3 View Figure 3 , 4 View Figure 4 ). The morphology of the maxilla is rather difficult to recognize, often fragmented; the main axis of this bone appears to be slightly curved. The outer mandibular surface is sculptured, ornamented with delicate striae. The dentary is relatively large, with a distinct pointed symphysial spine; the alveolar process of this bone bears minute granular teeth similar to those of the premaxilla. The articular bears a stout coronoid process and a rounded posteroventral bony lamina. The angular is poorly preserved in the material examined.
The suspensorium ( Figs 3 View Figure 3 , 4 View Figure 4 ) consists of the ectopterygoid, endopterygoid, hyomandibula, metapterygoid, palatine, quadrate, and symplectic. The hyomandibula is forked dorsally, producing two articular heads; the main shaft of this bone is stout; a short opercular process arises along the upper sector of the posterior margin. The quadrate is approximately triangular, with a wide dorsal head. The structure of the symplectic is not clear. The metapterygoid appears to be subrectangular in shape. The anterior portion of the palatine is curved and expanded; there is no evidence of palatine teeth or a dorsal articular head. The ectopterygoid is thick, wellossified, and expanded posteroventrally.
The bones of the opercular series ( Figs 3 View Figure 3 , 4 View Figure 4 ) are, in many cases, extensively fragmented because of their delicate, papery structure. The preopercle is crescentshaped. The opercle consists of a number of bony striae that radiate from the anterior condyle. The exact structure of the subopercle is difficult to determine.
The hyoid bar ( Figs 3 View Figure 3 , 4 View Figure 4 ) is robust and strongly elongate. The anterior ceratohyal is subrectangular, with a central constricted area. The posterior ceratohyal is nearly triangular in outline. There are five, slender, pointed branchiostegal rays.
The bones of the branchial arches are often fragmented. The ceratobranchials are slender and curved medially ( Fig. 4 View Figure 4 ). The epibranchials are thin and short; these bones can be observed exclusively in CM 5237. Sparse branchial teeth, possibly belonging to the second and third pharyngobranchials and fifth ceratobranchial can be also observed in some specimens ( MGPD 68487; MCZ 5177a, b).
The vertebral column ( Fig. 3 View Figure 3 ) is relatively linear and straight. There are 19 (or rarely 20) vertebrae, including the last half-centrum fused to the hypural plate. The centra are rectangular, slightly longer than high. The anterior-most vertebra is closely associated with the posterior margin of the neurocranium. The neural spines of the second to fifth anterior-most vertebrae are greatly expanded anteroposteriorly. The vertebrae bear prominent neural prezygapophyses. The abdominal vertebral centra (five) six to nine (ten) bear well-developed posteriorly directed haemal spines progressively increasing in size. The haemal spines of the caudal portion of the body are robust and slender. The neural and haemal spines of the penultimate centrum (second preural centrum) are spatulate and broadly expanded anteroposteriorly. The hypural plate is large, triangular in outline, with a short median notch on the posterior margin ( Fig. 3 View Figure 3 ). All values are in millimetres. Values in parentheses are in per cent of standard length.
There is no trace of an epural. There are nine (four + five) caudal-fin rays, all segmented and branched, except the ventral-most, which is reduced to a small unsegmented splint.
All three dorsal-fin spines are well developed ( Figs 3A View Figure 3 , 4 View Figure 4 ). The illicium and second dorsal-fin spine are situated anterodorsal to the neurocranium. The anterior-most (illicial) pterygiophore, which supports the illicium and second dorsal-fin spine, is hypertrophied, with a greatly enlarged, blade-like ventral margin; the anterior margin of this bone is irregular, characterized by a small process and two closely spaced foramina for the articulation of the two spines. The ventral blade-like bony lamina of this pterygiophore shows thick striations and alternating furrows that radiate approximately from the anterior margin of the bone. The illicial bone is slender and delicate, whereas the second and third dorsal-fin spines are elongate and relatively robust. The pterygiophore of the third dorsal-fin spine partially overlaps the posterior end of the preceding pterygiophore. The soft dorsal fin consists of 12–13 rays supported by 12 pterygiophores ( Fig. 3 View Figure 3 ). The pterygiophores are short, massive, and laterally compressed, with a proximal fan-like articular portion. Proximally, the posterior-most pterygiophore bears an enlarged rounded posterior process. The main axis of the five anterior-most pterygiophores of the soft dorsal fin is short and does not interdigitate with the neural spines of the underlying vertebrae. The anal fin is extremely short, containing eight or nine rays supported by seven pterygiophores ( Fig. 3 View Figure 3 ). All the pterygiophores are obliquely orientated, with their proximal portion directed posteriorly. The two posterior anal-fin pterygiophores insert between the haemal spines of the 14th and 15th vertebrae. Proximally, the posterior-most anal-fin pterygiophore bears an enlarged, rounded posterior process, similar to that of the opposite pterygiophore of the soft dorsal fin.
The pectoral fin originates just above the anal-fin origin and contains six or seven filamentous rays ( Fig. 3 View Figure 3 ). The extreme posterior displacement of the pectoral-fin origin is related to the enormous elongation of the two pectoral radials (as long as 41.4% SL). The post-temporal is difficult to recognize. The supracleithrum is a blade-like ovoid bone, with rounded margins. A large part of the medial surface of the supracleithrum is bound to the lateral surface of the dorsal arm of a large, crescent-shaped cleithrum. There is a single rod-like postcleithrum. The coracoid and scapula are laminar, laterally compressed. The pelvic fin consists of a single spine and five unbranched rays ( Figs 3 View Figure 3 , 4 View Figure 4 ). The basipterygium is stout, with an expanded articular head. The skin is naked.
Remarks: As discussed above, the relationships and familial placement of † Histionotophorus bassani were actively debated ( Woodward, 1901; Eastman, 1904, 1905; Gill, 1904; Regan, 1912; Le Danois, 1964; Rosen & Patterson, 1969) until Pietsch (1981) conclusively demonstrated its alignment with the Brachionichthyidae . Our morphological investigation revealed a number of characters that unquestionably support the independent status of this taxon. Yet, at the same time, † Histionotophorus bassani is easily distinguished from all other members of the family in the conspicuous development of the spinous dorsal fin and the soft rays of the dorsal, anal, and caudal fins, making it look superficially similar to certain genera of the family Callionymidae , for example, Bathycallionymus , Calliurichthys , Dactylopus , Diplogrammus , Foetorepus , Orbonymus , and Pseudocalliurichthys (see Nakabo, 1982).
| T |
Tavera, Department of Geology and Geophysics |
| CM |
Chongqing Museum |
| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Histionotophorus
| Carnevale, Giorgio & Pietsch, Theodore W. 2010 |
Histionotophorus Eastman, 1904: 32
| Eastman CR 1904: 32 |
| De Zigno A 1887: 31 |
| De Zigno A 1887: 31 |
| Diesing KM 1851: 80 |
Histionotophorus bassani
| Frickhinger KA 1991: 695 |
| Pietsch TW 1981: 416 |
| Rosen DE & Patterson C 1969: 442 |
| Le Danois Y 1964: 141 |
| Regan CT 1912: 283 |
| Eastman CR 1905: 30 |
| Eastman CR 1904: 33 |
Histiocephalus
| De Zigno A 1887: 31 |
| De Zigno A 1887: 31 |
Histiocephalus bassani
| Woodward AS 1901: 579 |
| De Zigno A 1887: 24 |