
Heterodeviata sinica, Song & Dong & Lu & Al-Farraj & Song & Hines & Luo, 2023
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlad044 |
|
DOI |
https://doi.org/10.5281/zenodo.8334785 |
|
persistent identifier |
https://treatment.plazi.org/id/03D68792-FFF4-FF8F-049A-3382FBC8889C |
|
treatment provided by |
Plazi |
|
scientific name |
Heterodeviata sinica |
| status |
sp. nov. |
Heterodeviata sinica View in CoL sp.nov.
Zoobank registration: rn:lsid:zoobank.org:act: A51495B7- 9B30-4EE6-B9B3-CB9B6560EFC6
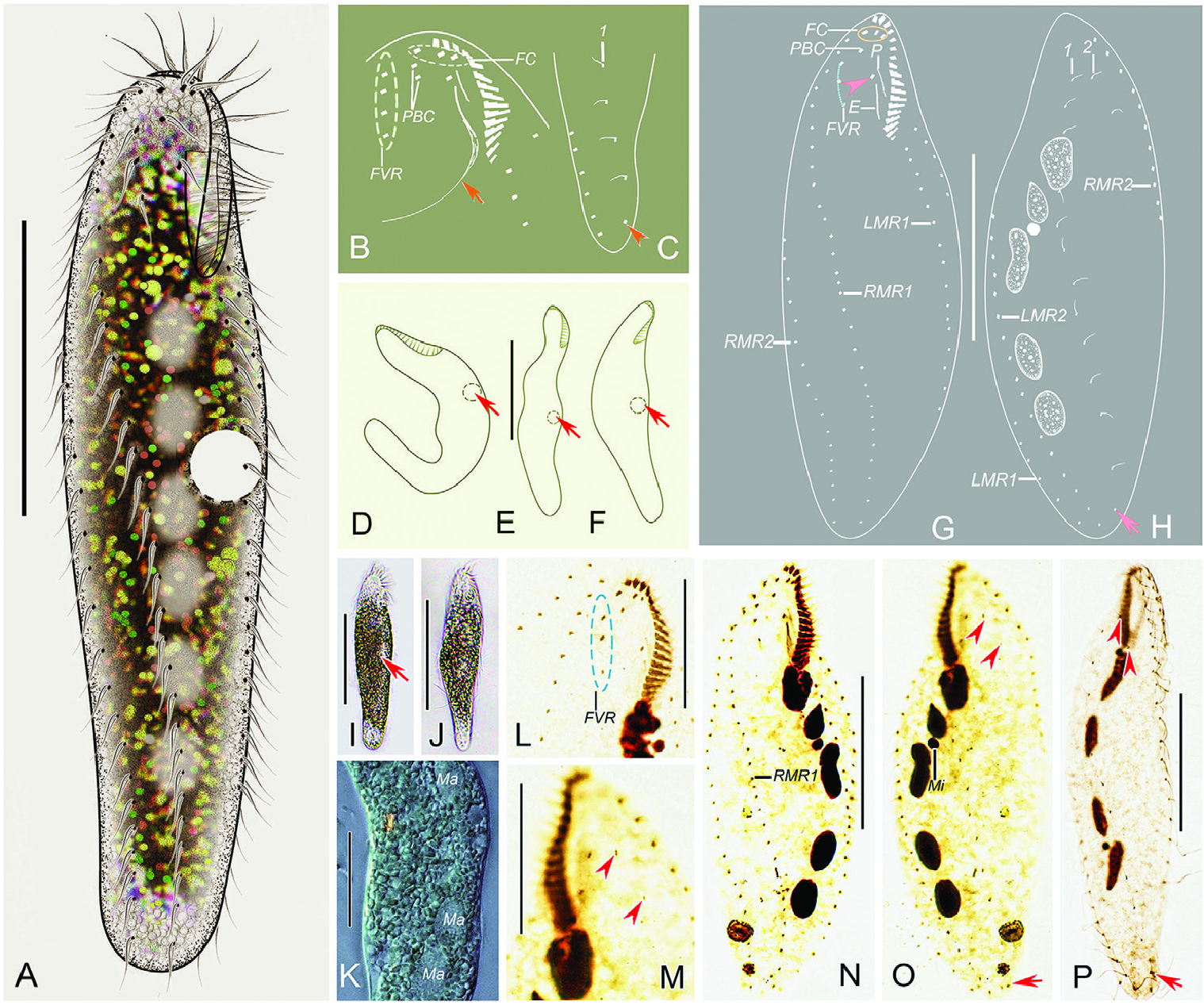
Diagnosis: Cell size 75–130 × 15–30 μm in vivo; elongated elliptical; contractile vacuole positioned at about 45% of cell length; adoral zone composed of about 16 membranelles; three frontal cirri, one buccal cirrus, one or two parabuccal cirri; one short frontoventral cirral row, two less and two right marginal rows; three to six macronuclear nodules; freshwater habitat.
Type material: A protargol slide (No. SWY2019091201/1) containing the holotype specimen, circled with black ink on the undersurface of the slide ( Fig.3G, H, N, O View Figure 3 ) and 12 paratype slides
(No. 2019091201/2–13) have been deposited in the Laboratory of Protozoology, Ocean University of China, ºingdao, China.
Type locality: Lake Weishan (34°44ʹ40ʹʹN, 117°09ʹ53ʹʹE), Shandong Province, China .
Etymology: The species-group name sinica (Latin adjective; Chinese or originating from China) means the species was first discovered in China.
Morphological description ( Fig.3A–P View Figure 3 ; Table 1)
Cell (freshly collected) size 75–130 × 15–30 µm (101 × 23 µm on average) in vivo (N = 8), ratio of length to width in vivo ranging 3.8–6.0:1 (on average 4.5:1); 86–138 × 20–63 μm (107 × 36 μm on average), with length: width ratio about 3.2:1 (range 2.2–4.7:1) in protargol-impregnated specimens. Cell flexible but not contractile, elongated elliptical, with both ends slightly narrowed and rounded ( Fig. 3A, D–F, I, J View Figure 3 ). Cortical granules absent. Cytoplasm colourless, filled with lipid inclusions with variable size, approximately 1–3 μm, rendering cells opaque except for the anterior and posterior portions of the cell, which appear transparent ( Fig. 3A, I–K View Figure 3 ). Three to six macronuclear nodules are situated in the less half of the cell, roughly ellipsoidal or dumbbell-shaped ( Fig. 3H, K, O, P View Figure 3 ). One to three oval micronuclei (about 3.2 μm in diameter, haphazardly placed touching macronuclear nodules) were observed in protargol preparations but difficult to recognize in vivo ( Fig. 3H, O, P View Figure 3 ). Contractile vacuole positioned close to less cell margin at about 45% of cell length, 10–14 μm across ( Fig. 3A, D–F, I View Figure 3 ). Collecting canals absent. Locomotion slow, usually gliding over substrate on surface of Petri dishes with cell obviously twisted. No cysts or conjugation were observed, although both are likely present within populations.
Adoralzoneofmembranelles (AZM) roughlyin Gonostomum - paưern, composed of 12–20 membranelles. Buccal field narrow and inconspicuous, occupies about 25% of cell length in vivo and on average 21% of cell length (range 17–26%) asser protargol impregnation ( Fig. 3A, D–F, G, N View Figure 3 ; Table 1). Paroral and endoral almost straight, nearly the same length and both single-rowed ( Fig. 3B, G, N View Figure 3 ). Pharyngeal fibres conspicuous in most stained specimens ( Fig. 3B View Figure 3 ). Cirri about 9–11 μm in vivo, constantly three slightly enlarged frontal cirri; one buccal cirrus located anterior of endoral ( Fig. 3B, G, N View Figure 3 ). One or two parabuccal cirri distributed behind right frontal cirrus ( Fig. 3B, G, N View Figure 3 ). One short frontoventral cirral row, with three to five (c. four) cirri, commences slightly behind right of parabuccal cirri and terminates before the proximal end of AZM ( Fig. 3B, G, L, N View Figure 3 ). Two less marginal rows (LMR), LMR1 (the inner row) and LMR2 with 17–26 and 18–38 cirri, respectively, both begin close to the rear end of the adoral zone ventrally, then usually extend on to dorsolateral side posteriorly ( Fig. 3G, H, N, O View Figure 3 ). Two right marginal rows (RMR), RMR1 with 19–43 cirri, begins at the right side of the short frontoventral cirral row and terminates near the end of the cell, RMR2 with 17–40 cirri, commences nearly the anterior end of cell on the dorsal side and extends to the ventral side at about two-fisshs of cell length ( Fig. 3G, H, N, O View Figure 3 ).
Two dorsal kineties (DK), the dorsal bristles inconspicuous in vivo, about 2.5 μm in protargol-impregnated specimens. DK1 with 10–20 dikinetids, DK2 with constantly two dikinetids. One caudal cirrus at the end of DK1 ( Fig. 3C, H, M, O, P View Figure 3 ).
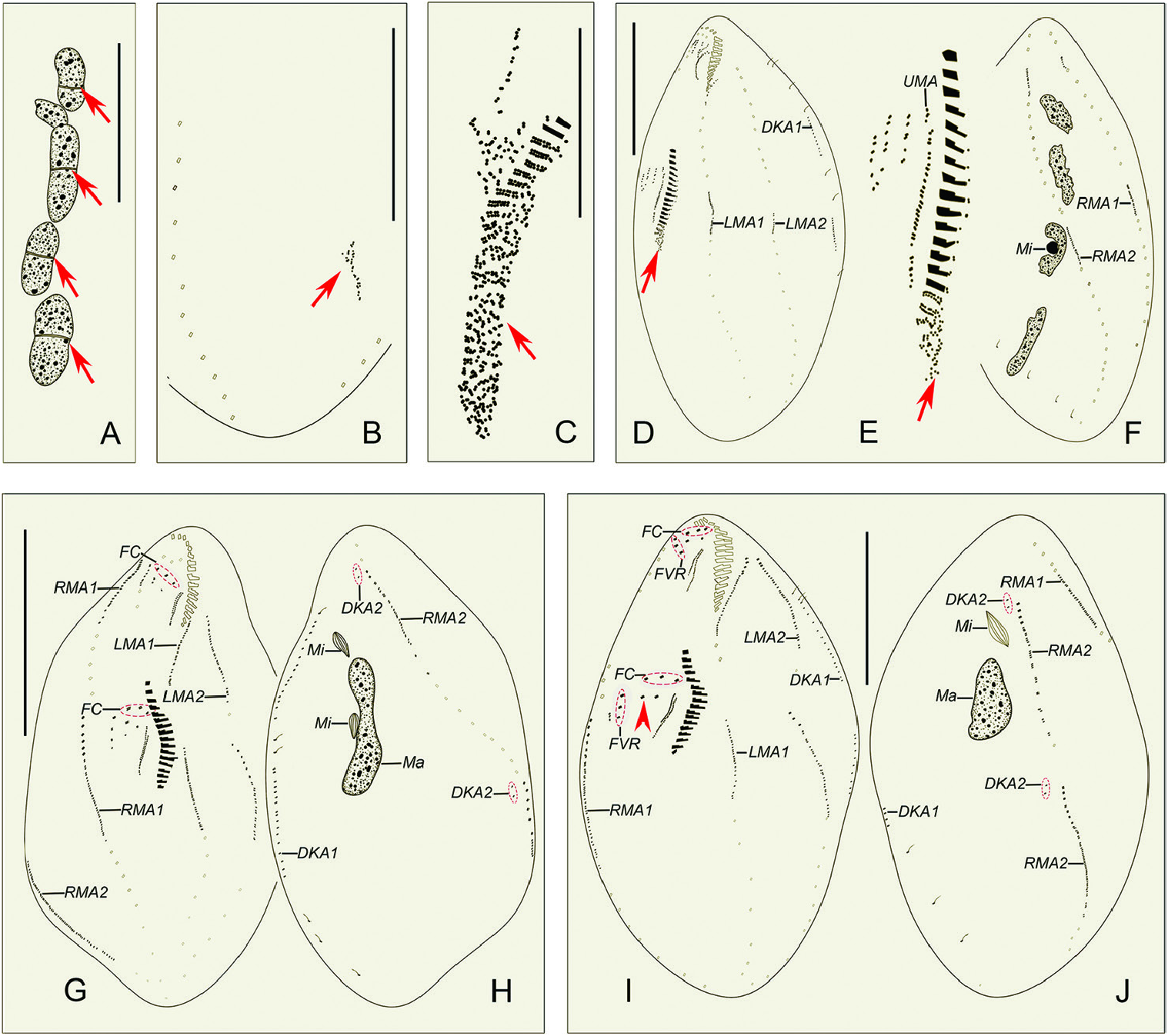
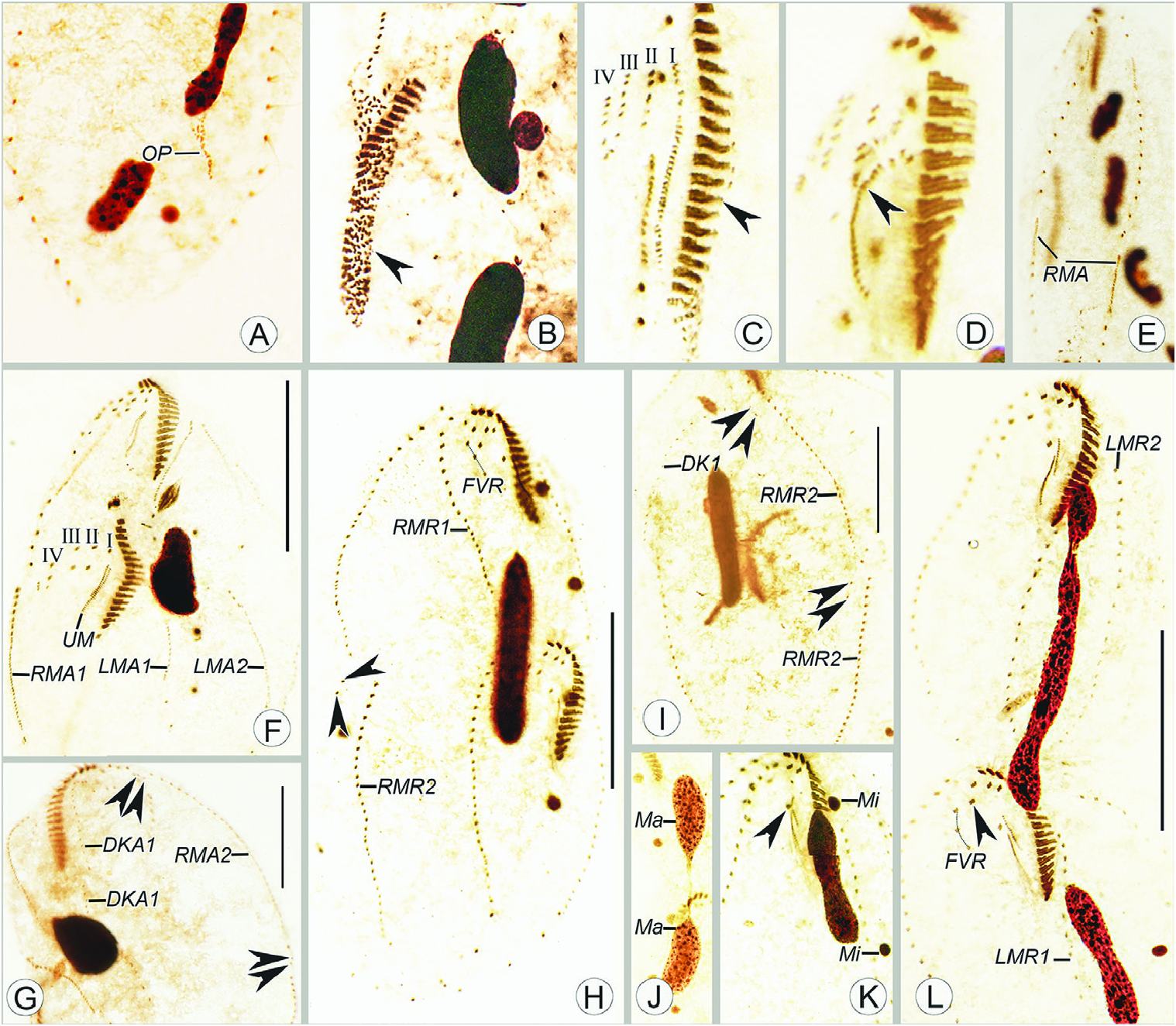
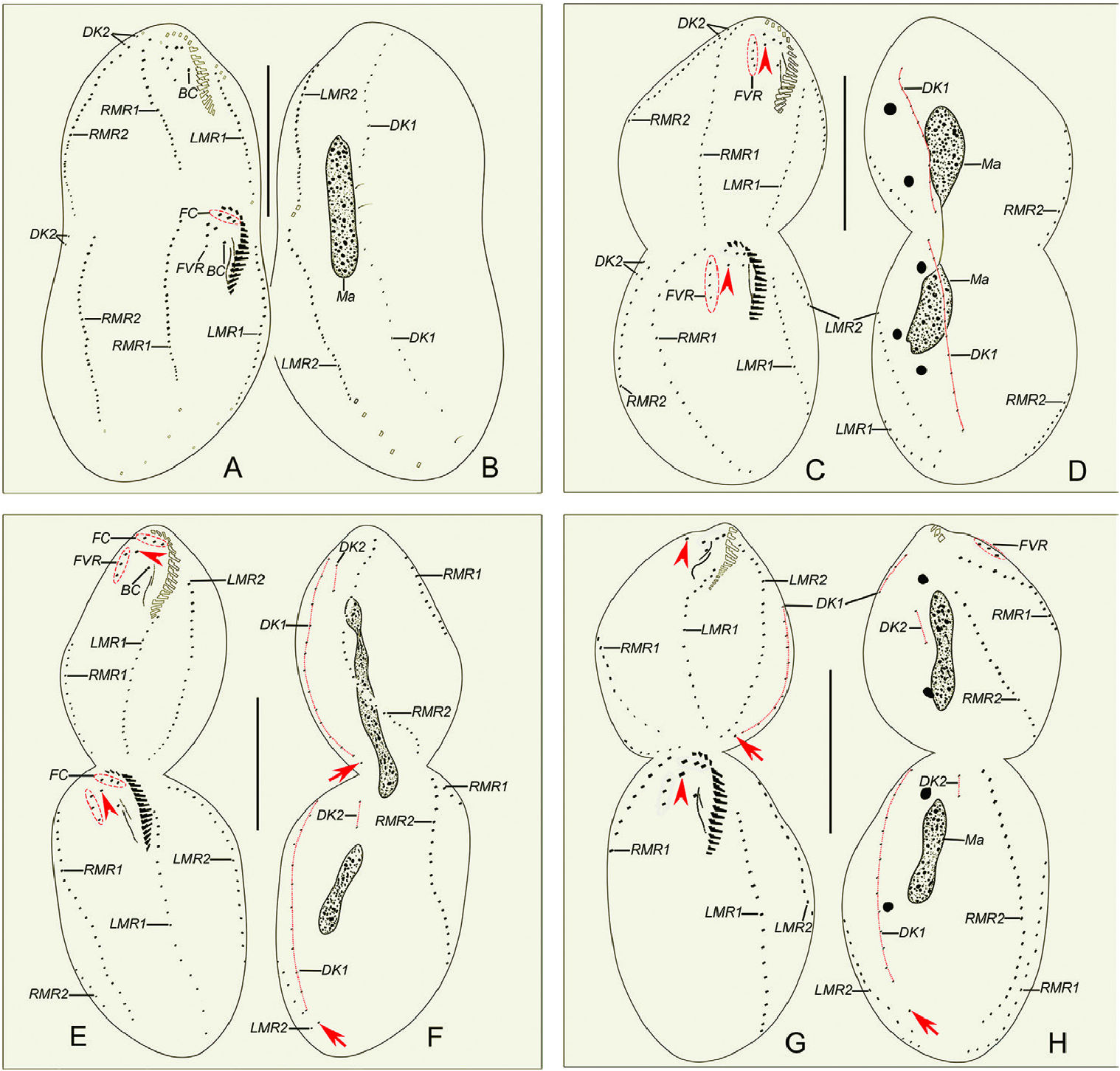
Morphogenesis ( Figs 4A–J View Figure 4 , 5A–L View Figure 5 , 6A–H View Figure 6 )
The following morphogenetic features are based on 10 dividers observed in the protargol preparations. No conjugation was observed.
Stomatogenesis, flontoventral cirri, and marginal rows
In early dividers, the oral primordium of the opisthe develops de novo in the posterior part of the cell between the inner less and right marginal row ( Figs 4B View Figure 4 , 5A View Figure 5 ). Subsequently, the oral primordium enlarges by further proliferation of basal bodies, several membranelles form in the anterior portion, and undulating membranes anlage (UMA = anlage I) appears in the opisthe ( Figs 4C View Figure 4 , 5B View Figure 5 ). Then, for the opisthe, another three cirral anlagen (anlagen II–IV) appear to the right of UMA ( Figs 4D, E View Figure 4 , 5C View Figure 5 ). At the same time, the parental endoral begins to disintegrate and anlagen I–IV also appear for the proter ( Figs 4D View Figure 4 , 5D View Figure 5 ). However, due to the lack of observations for some key division stages, it is unclear whether the old parental cirri contribute to the construction of the anlagen. In earlymiddle dividers, the formation of the oral primordium for the opisthe is gradually completed from anterior to posterior, and the UMA (anlage I) for each daughter cell splits longitudinally into undulating membranes and gives rise to the less frontal cirrus ( Fig. 4G View Figure 4 ). In middle to late dividers, cirral anlagen II–IV for the proter and opisthe differentiate into new cirri in the following paưern: anlage II forms the middle frontal cirrus and the buccal cirrus; anlage III forms the right frontal cirrus and one or two parabuccal cirri; anlage IV forms three to five ventral cirri ( Figs 4I View Figure 4 , 5F, H, K, L View Figure 5 , 6A, C, E, G View Figure 6 ).
Two less marginal anlagen (LMA) for the opisthe develop in the middle of the parental structures ( Fig. 4D View Figure 4 ). Meanwhile, two right marginal anlagen (RMA) for both proter and opisthe form intrakinetally ( Figs 4F View Figure 4 , 5E View Figure 5 ). In the next stage, LMA for the proter develop in the anterior part of the parental structures ( Fig. 4G View Figure 4 ). From middle to late stages, both less and right marginal anlagen extend gradually bidirectionally and, finally, the marginal cirral anlagen begin to differentiate into new cirri for the proter and opisthe and the parental marginal cirri are resorbed ( Figs 4I, J View Figure 4 , 5F–I, L View Figure 5 , 6A–H View Figure 6 ).
Dorsal ciliature
In early dividers, two thread-like dorsal kineties anlagen (DKA1), one for proter and another for opisthe form within the parental DK1 ( Fig. 4D View Figure 4 ). In middle to late dividers, DKA1 (dorsomarginal kinety) develops with the proliferation of basal bodies, and DKA2 appears to the right of the new right marginal row 2 for both proter and opisthe ( Figs 4H, I, J View Figure 4 , 5G View Figure 5 ), then they give rise to the new DK1, 2 ( Figs 5H, I View Figure 5 , 6A–D, F, G, H View Figure 6 ). During the morphogenetic process, one caudal cirrus is formed at the end of DK 1 in both the proter and opisthe ( Fig. 6F, H View Figure 6 ).
Nuclear apparatus
In the very early stage, a replication band is present in each macronuclear nodule ( Fig. 4A View Figure 4 ). The macronuclear apparatus divides in the usual way for hypotrichs, i.e. the macronuclear nodules fuse into a single mass during middle stages, which then splits two or three times prior to cytokinesis, and new macronuclear nodules are formed for each daughter cell ( Figs 4F, H, J View Figure 4 , 5F–H, J–L View Figure 5 , 6B, D, F, H View Figure 6 ). The micronuclei divide mitotically ( Figs 4F View Figure 4 , 5K, H, J View Figure 5 , 6D, H View Figure 6 ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.