
Guibemantis woosteri, Lehtinen & Glaw & Vences & Rakotoarison & Scherz, 2018
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.451 |
|
publication LSID |
lsid:zoobank.org:pub:40E9A16C-69E9-47FA-8588-A9F2FFFB63D9 |
|
DOI |
https://doi.org/10.5281/zenodo.3815614 |
|
persistent identifier |
https://treatment.plazi.org/id/BB4E147F-EB7A-41F9-9E1B-94B92521EE7F |
|
taxon LSID |
lsid:zoobank.org:act:BB4E147F-EB7A-41F9-9E1B-94B92521EE7F |
|
treatment provided by |
Valdenar |
|
scientific name |
Guibemantis woosteri |
| status |
sp. nov. |
Guibemantis woosteri View in CoL sp. nov.
urn:lsid:zoobank.org:act:BB4E147F-EB7A-41F9-9E1B-94B92521EE7F
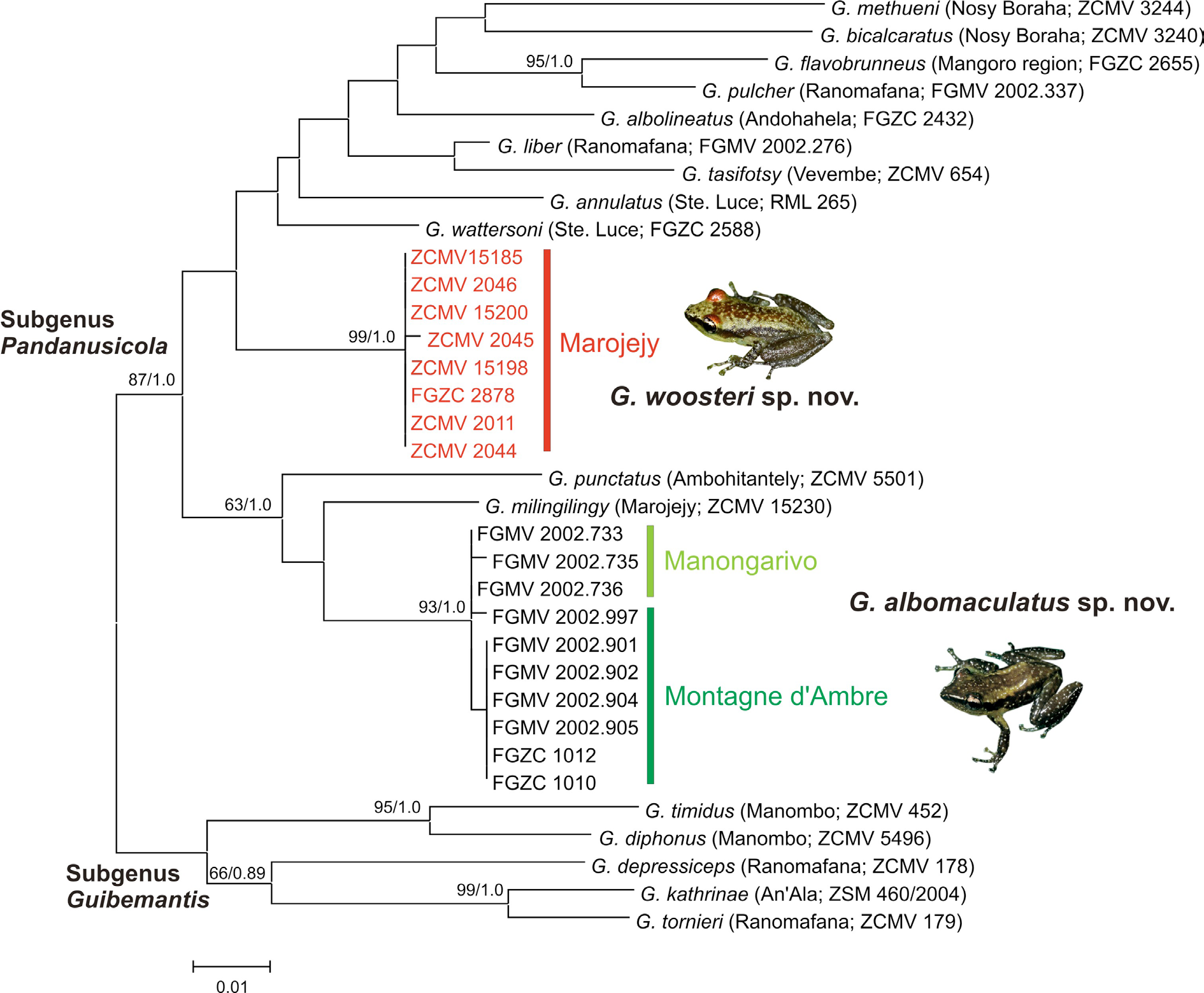
Figs 1–3 View Fig View Fig View Fig , 5 View Fig
Suggested common name: Wooster’s Malagasy Pandanus Frog.
Diagnosis
Assigned to the subgenus Pandanusicola of the genus Guibemantis based on its small body size, phytotelm-breeding habitats (in Pandanus plants), moderate webbing between toes, connected lateral metatarsalia, the presence of both inner and outer metatarsal tubercles, type 2 femoral glands in males, and DNA sequence similarities ( Glaw & Vences 2006). The new species is characterized by the unique combination of the following characters: (1) male SVL 19.4–21.8 mm, (2) two fairly conspicuous parallel light-colored dorsolateral lines on the dorsum, (3) well-defined dark bars on the forearms and sometimes on the legs, (4) a conspicuous light area near the forearm insertion, (5) dark ventral surfaces to hands and feet, (6) thick, single dark rostral stripe, (7) distinctly reddish color dorsally surrounding the eyes, (8) white spots of the dorsum and limbs and (9) gray femoral glands in males.
Etymology
This new species is dedicated to the College of Wooster and its faculty, staff, students and alumni in celebration of 150 years of educational excellence.
Material examined
Holotype
MADAGASCAR: adult ♀, Marojejy National Park at “Camp Simpona”, 14.437° S, 49.743° E, 1326 m a.s.l., 18 Feb. 2005, F. Glaw, M. Vences and R.D. Randrianiaina leg. [ ZSM 5063 View Materials /2005 ( ZCMV 2044 )] ( Fig. 3 View Fig ).
GoogleMapsParatypes (n = 15)
MADAGASCAR: 8 adults, Manantenina River in Marojejy National Park, 700–1300 m a.s.l., Nov. 1992, R.A. Nussbaum, C.J. Raxworthy, A. Razafimanantsoa and A. Razafimanantsoa leg. ( UMMZ 212601– 212602, UMMZ 212604–212606, UMMZ 212613, UMMZ 212615, UMMZ 212617); adult ³, same data as for holotype [ ZSM 5062/2005 ( ZCMV 2011)]; 1 ♀, Marojejy National Park, Camp 3 “Simpona”, 14.4366° S, 49.7434° E, 1325 m a.s.l., 17 Nov. 2017, M.D. Scherz, A. Rakotoarison, M. Bletz, M. Vences and J. Razafindraibe leg. [ ZSM 425/2016 ( ZCMV 15185)]; 1 ³ and 1 juv., Marojejy, between Camp 2 “Marojejia” and Camp 3 “Simpona”, no coordinates taken, 18 Nov. 2016, M.D. Scherz, A. Rakotoarison, M. Bletz, M. Vences and J. Razafindraibe leg. [ ZSM 426/2016 ( ZCMV 15198), UADBA uncatalogued ( ZCMV 15200)]; 1 spec., Marojejy National Park, same data as for holotype, but collected on 16 Feb. 2005 [( FGZC 2878), UADBA uncatalogued]; 1 spec., same data as holotype ( ZCMV 2045); 1 spec., same data as holotype, but collected above Camp Marojejia ( ZCMV 2046).
The paratypes from UADBA were not available for morphometric comparison, but were sequenced and attributed genetically to G. woosteri sp. nov. (see Fig. 1 View Fig ).
Referred specimens
Based on similarity in coloration (no genetic data available), we also tentatively assign to this species the series UMMZ 191339–191340 and UMMZ 191343–191344; adults from Ambatovaky Special Reserve (600–800 m), collected by C.J. Raxworthy on 21 February 1990.
Description of the holotype
Adult female. Head longer than wide and slightly wider than body; snout rounded in dorsal, ventral and lateral views; canthus rostralis rounded; nostrils very near to the tip of snout, internarial distance 79% of interorbital distance; tympanum distinct 38% of horizontal eye diameter; very dark and prominent supratympanic fold present from posterior edge of eye around the tympanum nearly to the arm insertion; one small round patch of vomerine teeth medial between eye and choanae on either side of head; tongue stout and strongly bifid. Arms thin, lower arm is 93% of hand length; relative finger length 1<2<4<3, finger discs moderately enlarged and squared off at tips in a rounded ‘T’ shape, only traces of webbing between fingers, subarticular tubercles prominent. Hindlimbs relatively thin, femur length 91% of tibia length, foot length 88% of tibia length; lateral metatarsalia connected; inner metatarsal tubercle oblong, (0.8 in length, 0.3 in width); outer metatarsal tubercle round (0.3 in diameter); webbing formula between toes 1(1) 2i(1) 2e(1) 3i(2) 3e(2) 4i(2.75) 4e(3) 5(1.5); relative length of toes 1<2<5<3<4; toe discs moderately enlarged, width of fourth toe disc 92% of width of third finger disc. Cloaca not clearly recognizable; skin is finely granular dorsally, more coarsely granular ventrally. For morphometric measurements see Table 1. A View Table 1 sample of muscle was taken from right leg for genetic analysis.
After 13 years in preservative, the dorsal background coloration is a medium brown with the flanks much darker brown. Starting at the snout tip, there is a distinct color change along the dorsolateral area (medium brown medially, dark brown laterally). A few vague light spots are present, but there are no conspicuous dorsal markings. A thick, dark rostral stripe is present. The dorsal surface of arms, hands and fingers is medium brown with darker bands on the forearms. Dorsal surface of legs, feet and toes is a uniform medium brown with no obvious markings. The skin on the upper arms near the articulation with the body conspicuously lacks any coloration and this contrasts markedly with the more distal portions of the arm that are darker. Finger discs are fairly dark brown dorsally and ventrally with some lighter areas (especially fingers I and II). Toe discs are similar. Fairly obvious white annuli are present on many digits. Ventral surface cream with dark spots on jawline and ventral surface of thighs, hands and feet.
Coloration in life
The life coloration of the holotype is unknown. However, based on photographs of live individuals from Marojejy ( Fig. 5 View Fig ), there are several clear coloration differences compared to preserved specimens. The small white spots, often prominent in life, are not usually obvious after preservation in alcohol. The iris, which is gold in life, is white or gray in preservative. The portions of the limbs, dorsum and head, medium brown in preservative, appear yellowish-green in life. Moreover, at least some individuals have a prominent red color on the eye orbits; there is no indication of this in preservative. The ventral skin is semi-translucent in life such that chest and belly musculature are visible (not apparent in preservative). In males, femoral glands (type 2, as defined by Glaw et al. 2000) appear gray in life with large granules in the gland, each of which has a white spot ( Fig. 5C View Fig ). Sexually mature males also possess a white coloration on portions of the throat.
Variation
Like most Pandanusicola , this new species is small (overall mean 22.1 ± 1.6 mm SVL, n = 14). Females are larger (mean 23.8 ± 1.0 mm SVL, n = 5) than males (mean 21.1 ± 0.8 mm SVL, n = 9) on average. Some individuals have relatively few small white dorsal spots (primarily on the hindlimbs), while others have dense spots over much of the dorsal surfaces (contrast Fig. 5A and 5B View Fig ). The light dorsolateral lines on the dorsum are conspicuous in some individuals but much less so in others. Dark spotting on the dorsum also varies from none to many. Dark bars on legs vary from reasonably distinct ( Fig. 5A View Fig ) to inconspicuous ( Fig. 5B View Fig ). Head coloration can be variable also, with several specimens having a distinctly lighter coloration anterior to the eyes than posterior and one specimen possessed a distinct interocular bar. Many, but not all, specimens had a distinct light spot in the middle of the snout. The color of the digits and toepads also varies. Some individuals have distinct dark bars on the digits and this seems to be an extension of the bars on the forearms. The color of the toepads is similarly variable with the pads on some digits being dark and other digits being light. A few individuals had indications of faint annuli on the digits proximal to the toepads or a second rostral line parallel to the first (as in G. annulatus and G. wattersoni ), however, these are usually not well developed. Ventrally, coloration varies from having dark markings only along the jawline to individuals with dark spotting on the throat, chest and/or thighs. The ventral surfaces of the hands and feet are apparently always dark. One individual (UMMZ 212604) had especially rough granular patches on the belly, reminiscent of those recently described from G. tasifotsy . No variation was observed in the webbing on the hand. However, webbing on the foot was variable for some digits (range, where variable, given in parentheses): 1(1) 2i(1) 2e(1) 3i(2) 3e(1–2) 4i(2.5–3) 4e(2.5–3) 5(1–2).
Differential diagnosis
Among Pandanusicola , the new species is distinguished from G. annulatus , G. bicalcaratus , G. flavobrunneus , G. liber , G. pulcher , G. punctatus and G. tasifotsy by the presence of two conspicuous parallel dorsolateral light-colored lines on the dorsum, by the presence of small white spots on more darkly colored arms, legs and flanks and by large genetic differences. By comparison, G. pulcher is a characteristic bright green in color with large irregular dark dorsal spots; G. bicalcaratus , is yellowish or brownish dorsally with usually relatively few irregularly shaped dark spots; G. flavobrunneus is much larger (up to 38 mm SVL; Glaw & Vences 2007) and is primarily yellow with extensive brown dorsal markings; G. liber is rather variable in coloration but tends to be brownish overall, the entire surface of the throat of males is bright white, and this species does not breed in Pandanus plants; G. tasifotsy does not breed in Pandanus either, and has a copper metallic coloration on the nares and/ or eye orbits and prominent white lateral blotches; G. annulatus and G. punctatus are cream or olivecolored, respectively, and have abundant small, well-defined dark spots; G. methueni and G. wattersoni can have parallel light-colored dorsolateral lines on the dorsum but most individuals lack this feature and, if present, it is usually poorly developed. Furthermore, G. wattersoni is a yellow or light brownish frog dorsally and its flanks and legs are always lightly colored whereas they are typically dark in the new species. In G. methueni , the ventral surfaces of the hands and feet are very lightly colored, whereas they are dark in the new species. Guibemantis albolineatus lacks both vomerine teeth and a rostral stripe (both present in the new species), has a distinctly shaped snout that is square in dorsal and ventral view and is notably smaller in overall body size (see Table 1 View Table 1 ). The recently described G. milingilingy differs by lacking the characteristic white spots on the dorsum and limbs (versus presence), by lacking the reddish colored skin on the eye orbits (versus presence) and by having yellow femoral glands in mature males (gray in the new species). Finally, G. woosteri sp. nov. differs from G. albomaculatus sp. nov. in having well-defined dark bars on the forearms and sometimes on the legs, a conspicuous light area near the forearm insertion (both of which G. albomaculatus sp. nov. lacks) and gray femoral glands in males with distinct white spots (femoral glands are orange in male G. albomaculatus sp. nov.). No other described species of Pandanusicola has the combination of light colored parallel dorsolateral lines with small white spots on the legs and flanks and well-defined dark bars on the forearms.
Distribution
Currently, G. woosteri sp. nov. is reliably known only from mid-elevation rainforests in Marojejy National Park, the type locality, in the North-East of Madagascar ( Fig. 2 View Fig ), from roughly 700–1325 m a.s.l. In addition, we here consider specimens from the Ambatovaky Special Reserve in the Northern Central East of the island as possibly belonging to this species ( Fig. 2 View Fig ), but this identification requires confirmation by molecular data.
Based on the limited knowledge of the distribution, we suggest a conservation status of "Data Deficient".
Natural history
Only known from Pandanus plants. One adult female attributed to this species (UMMZ 212617) had four visible eggs in the body cavity (egg diameter 0.8 mm) and another (UMMZ 191340; from Ambatovaky) had six (egg diameter 1.8 mm) possibly suggesting a small clutch size. Tadpoles and breeding vocalizations are unknown.
Available names and comparisons
The only available synonyms that could possibly be applied to this species are Gephyromantis albogularis Guibé, 1947 and Gephyromantis variabilis Millot & Guibé, 1951 , both of which were confidently assigned to Guibemantis liber by Lehtinen et al. (2011). Guibemantis liber males have a throat that is bright white over its entire surface. The whitish coloration on the throat of males in G. woosteri sp. nov. does not cover the entire surface nor is it as brightly white as in G. liber .
Remark
In Vieites et al. (2009) and Perl et al. (2014), the holotype of G. woosteri sp. nov. (ZSM 5063/2005 / ZCMV 2044) and one paratype (ZSM 5062/2005 / ZCMV 2011) were listed as two different candidate species (Ca15 and Ca17, respectively). However, the 16S sequence data reported here show them to be nearly identical genetically ( Fig. 1 View Fig ) and they are also morphologically similar ( Table 1 View Table 1 ). Thus, we consider them to be conspecific. In another earlier molecular analysis, Lehtinen et al. (2007) found the holotype of G. woosteri sp. nov. (ZSM 5063/2005) to be phylogenetically related to UMMZ 212597 (also from Marojejy) and UMMZ 212922 from Cap Est (in the same region, but further east on the coast). However, these forms are 7.3% and 8.1% different in the 12S and 16S mitochondrial genes ( Lehtinen et al. 2007) and are morphologically distinct from G. woosteri sp. nov. We have, therefore, not included these specimens in the definition of G. woosteri sp. nov. as they likely represent additional undescribed taxa.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |