
Glyptobairdia trinodosa Teeter, 1975
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5175.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:44FB9C3D-3188-4BFB-BDB8-C1324729A396 |
|
DOI |
https://doi.org/10.5281/zenodo.7003528 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE6B50-FFD2-FF83-ECD6-A9EF6B641DBF |
|
treatment provided by |
Plazi |
|
scientific name |
Glyptobairdia trinodosa Teeter, 1975 |
| status |
|
Glyptobairdia trinodosa Teeter, 1975 View in CoL
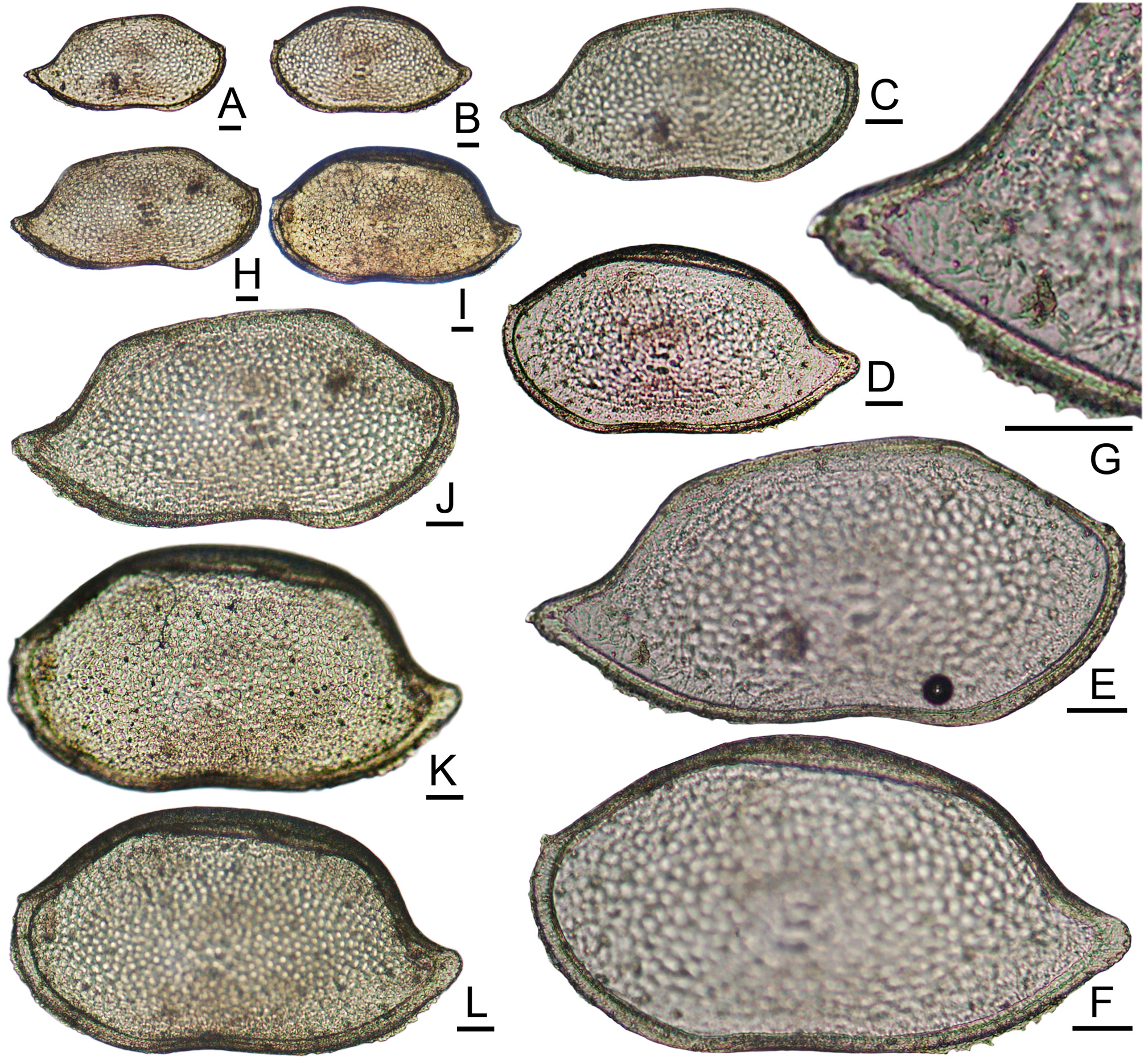
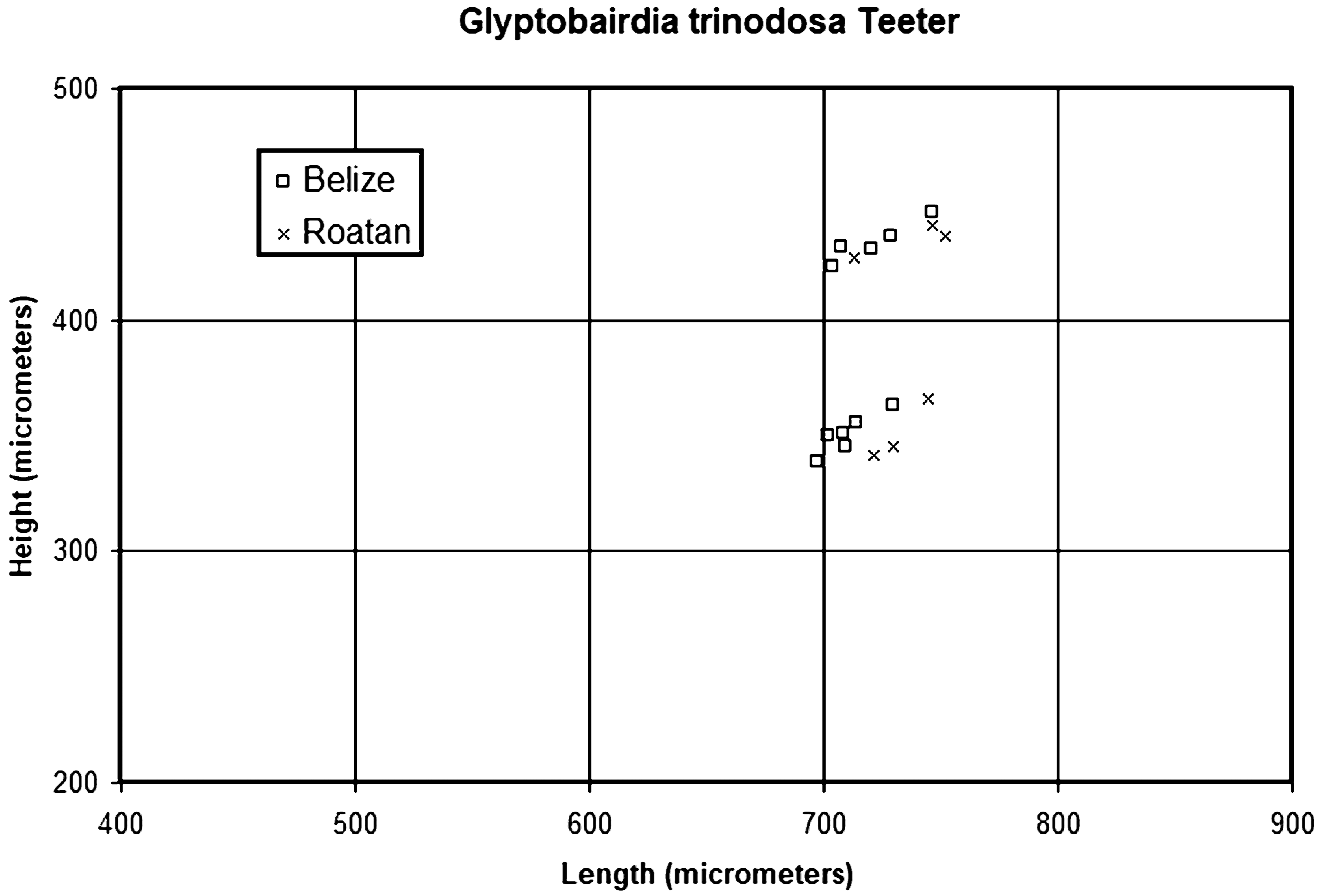
( Figs. 20 View FIGURE 20 , 21A–J View FIGURE 21 )
1975 Bairdoppilata (Glyptobairdia) trinodosa Teeter , n. sp.: p. 421, figs. 3h, 4f.
1983 Bairdoppilata (Glyptobairdia) binodosa [sic] Teeter.—Palacios-Fest et al., table 1.
1988A Glyptobairdia trinodosa Teeter. —Bold, table 2, p. 150.
Material: Sixteen subfossil carapaces and valves from near-reef sediments of the carbonate platform of Belize and Honduras.
Dimensions: Dry carapace specimen 4083W, LVL 0.720 mm, LVH 0.431 mm, RVL 0.708 mm, RVH 0.351 mm. Teeter reported carapace dimensions of L 0.70 mm, H 0.40 mm.
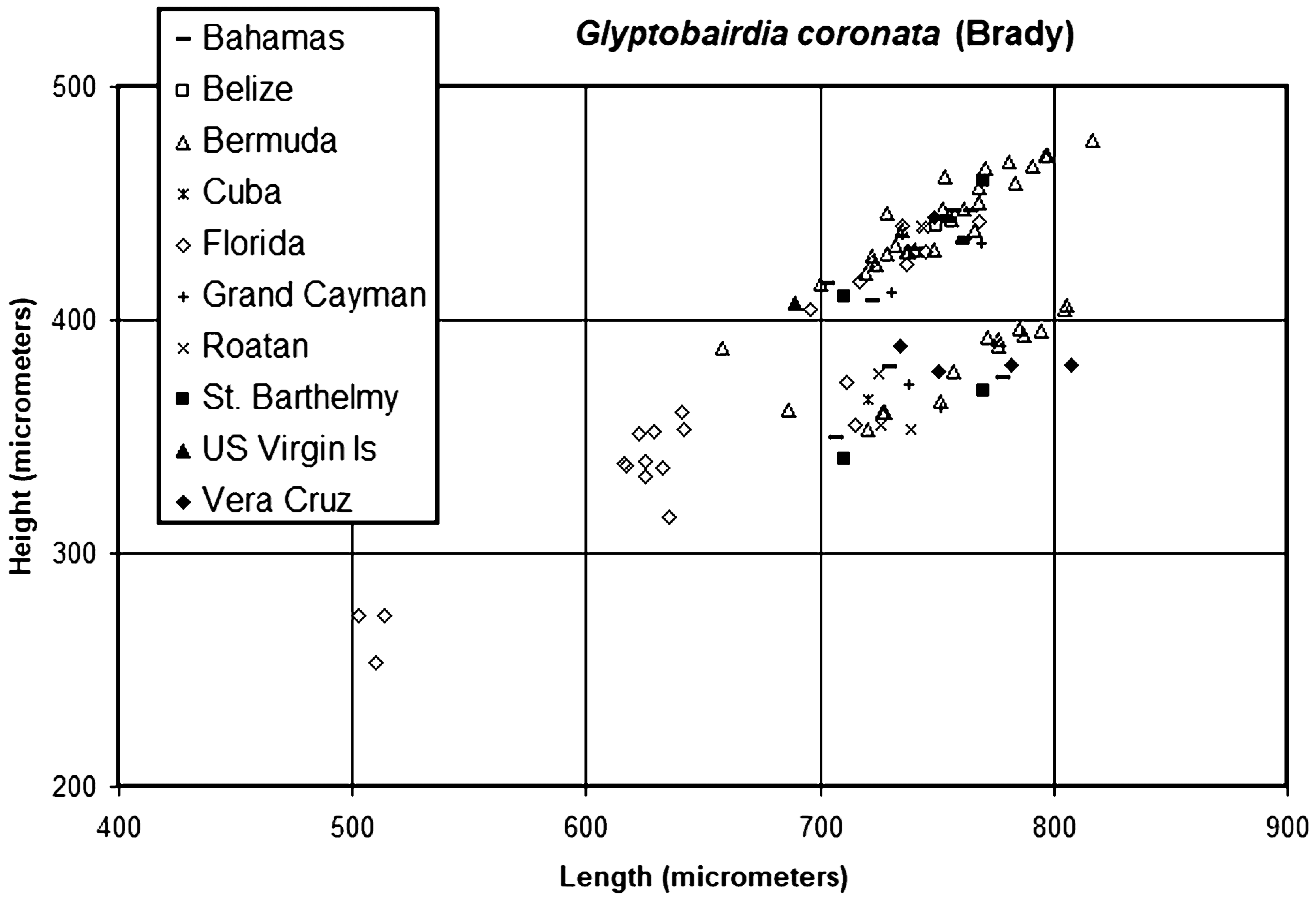
The H:L clusters for G. trinodosa ( Fig. 21 View FIGURE 21 ) are more compact than those for G. coronata , in part because fewer specimens were recovered from fewer localities. It is likely that females are longer and higher than males, but without soft parts this cannot be confirmed. The clusters of G. trinodosa ( Fig. 20 View FIGURE 20 ) overlap with the lower and middle parts of the clusters for G. coronata ( Fig. 17 View FIGURE 17 ), approximately equivalent to the Florida population of that species.
Anatomical Remarks: The subfossil valves are colorless, transparent to translucent, with a sugary texture and no opaque pattern. The two dorsal ends of the U-shaped central ridges on the lateral surfaces are thickened or flare into sharp crests ( Figs. 21A–H View FIGURE 21 ). A large round node is isolated a little above the gap between these ends. A second node is located in the middle of the central space, and the third node is positioned anteroventrally at a little distance from the second. This ornamental configuration is easily distinguished from that of G. coronata .
Teeter mentioned but did not illustrate supposed sexual dimorphism: “Dimorphism present in left valve; posteroventral part of submarginal rim depressed in males, continuous in females.” This feature could not be confirmed in the specimens examined for this report. The “submarginal rim” mentioned by Teeter actually consists of two low, rounded, crescentic (concave-up) folds in the marginal fabric, located in the anteroventral and posteroventral regions. They more or less coincide with the valve margins as seen in lateral view, but not the valve edges. They do not meet at midlength, nor do they extend dorsally. Because of the rather coarse pattern of punctation, which crosses these ridges, small nicks or breaks in the narrow muri may appear to interrupt the ridges in some specimens. These are accidental features, probably post-mortem.
Like that of G. coronata , the RV hinge has prominent, crescentic terminal teeth, which are directed laterally (into corresponding sockets of the LV) and project dorsally above the level of the median hinge-bar ( Fig. 21I View FIGURE 21 ). This is a contrast to the hinge of most bairdioids, in which the ends of the RV hinge-bar are less elevated relative to the median bar and less tooth-like in form. The hinge elements of G. trinodosa display coarse ligamental crenulation, like that of G. coronata ( Figs. 21I–J View FIGURE 21 ).
The soft parts and esophageal valve of this species are unknown.
Taxonomic Remarks: Bold (1974, p. 34) initially suggested that “this may be no more than a local variant” of G. coronata , but the differences between these two species are consistent, and specimens are easily sorted where they occur together.
On the Belize platform, Teeter (1975, Table 1 View TABLE 1 ) reported nine adult specimens plus uncounted juveniles of G. trinodosa at six collecting stations. For G. coronata , he collected eight specimens at four stations, and the two species occurred together in one sample. The occasional co-occurrence of these two uncommon species is confirmed in the near-reef assemblages of Belize and Honduras examined for this report .
Teeter (p. 421) stated that “The presumed juveniles of either this species or B. (G.) coronata indicate a remarkable ontogeny. They have the bairdian outline, show pronounced dorsal overlap, and have a punctate surface; but they lack any indication of the prominent ridges common to the adult.” The “dorsal overlap” he mentioned probably refers to overreach. In his Table 1 View TABLE 1 , juvenile specimens were recorded at just one station and identified as G. trinodosa , without explanation. Illustrations and dimensions were not provided for the juveniles. No adults of either species were recorded in that sample, and no juveniles of G. coronata were tabulated at any station.
Here, juveniles of G. coronata from collections in the Florida Keys are illustrated for the first time ( Figs. 19 View FIGURE 19 AL). In the subfossil assemblages from Belize and Roatan examined for this report, no juveniles of either species have yet been seen.
Geographic Distribution: G. trinodosa occurs consistently, though never abundantly, in near-reef assemblages of the Belize carbonate platform and Roatan Island (Bay Islands, Honduras). It has been reported as far north as Cozumel on the eastern shelf of the Yucatan platform (Bold 1988A, table 2, p. 150; Palacios-Fest et al. 1983).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Bairdioidea |
|
Family |
|
|
Genus |