
Gephyromantis verrucosus Angel, 1930
|
publication ID |
https://doi.org/ 10.5281/zenodo.212849 |
|
DOI |
https://doi.org/10.5281/zenodo.5672706 |
|
persistent identifier |
https://treatment.plazi.org/id/28038792-4C37-FFE7-FF7E-FD20314A7EE0 |
|
treatment provided by |
Plazi |
|
scientific name |
Gephyromantis verrucosus Angel, 1930 |
| status |
|
Gephyromantis verrucosus Angel, 1930 View in CoL
Resurrection, identity and lectotype designation. Gephyromantis verrucosus , considered a synonym of G. boulengeri for more than 60 years, is hereby resurrected and considered to represent a valid species. The two syntypes of Gephyromantis verrucosus (MNHN 1930.443 and 444) from a forest near Fort Carnot (at 600 m a.s.l.), Farafangana province (= Ikongo), are adult males, as recognizable from the presence of femoral glands in both specimens, and black color laterally on throat, representing vocal sacs, in MNHN 1930.444. Since this latter specimen is larger, in better state of preservation, and the secondary sexual characters are more clearly developed, we hereby designate this specimen as lectotype ( Fig. 12 View FIGURE 12 ).
The lectotype has distinct and large dorsal tubercle, arranged in very irregular rows and especially prominent dorsolaterally. In the shoulder region, the tubercles very weakly are arranged along an imagined line representing the inner dorsolateral ridges, but no such ridges can be reliably identified. Although color and pattern are largely faded on dorsal and ventral side (with some dark brown pigment remaining especially on the tubercles of the dorsolateral region), it is possible to recognize on the left side of the head clear dark and light alternating patches on the upper lip with two distinct dark patches interrupted by a light patch under the eye. No webbing is recognizable and the hands and feet are rather elongated and thin. Dark crossbands are recognizable dorsally on thigh, shank, and upper forelimb. The length and width of femoral glands are 4.2 x 1.9 mm in the lectotype and 3.2 x 1.8 mm in the paralectotype.
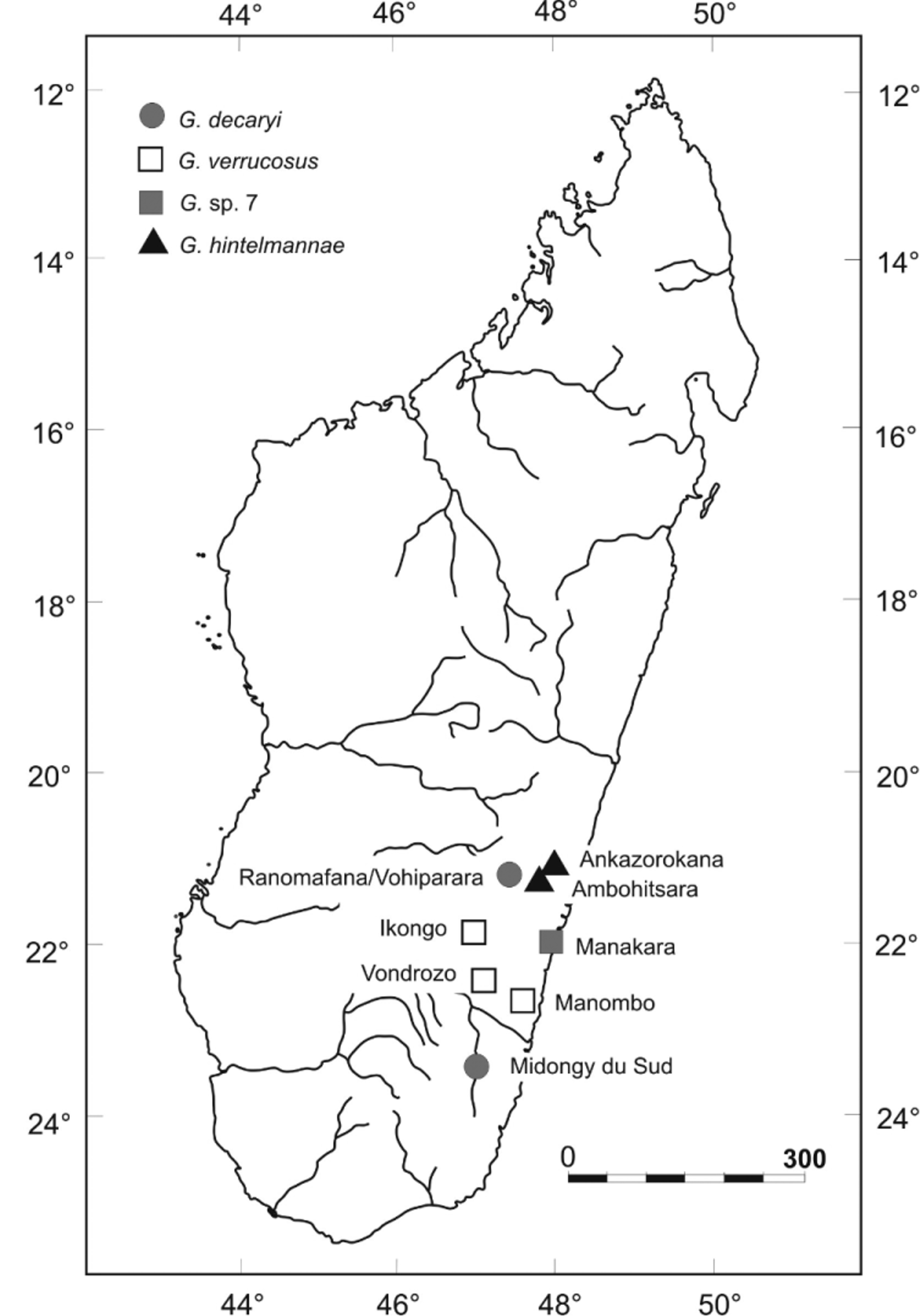
The lectotype thus agrees in most morphological and chromatic characters with the specimens collected by us at Manombo and Vondrozo. These localities are in the same general region of Madagascar's south-eastern coast as the G. verrucosus type locality Ikongo ( Fig. 10 View FIGURE 10 ). Consequently we here revalidate the name Gephyromantis verrucosus for these populations.
Remark. Vieites et al. (2009) and Wollenberg et al. (2011) considered the Vondrozo and Manombo populations as representing G. d e c a r y i, due to their geographical proximity to the type locality of decaryi (Midongy) . As argued above, we here instead follow Glaw & Vences (2000) and assign the name decaryi to the Ranomafana population. This species was referred to as G. sp. aff. decaryi (Vondrozo) by Kaffenberger et al. (2012).
Material examined. ZSM 2507/2007 ( ZCMV 5451), ZSM 2508/2007 ( ZCMV 5495), ZSM 2506/2007 ( ZCMV 5450), ZSM 2462/2007 ( ZCMV 5459), adult males, collected in Manombo, forest upstream, south-eastern Madagascar, coordinates not taken but less than 4 km from 23° 01.699' S, 47° 43.892' E, at ca. 50 m above sea level, by M. Vences, G. Safarek, E. Rajeriarison and T. Rajoafiarison on 24 February 2007; ZSM 2505/2007 ( ZCMV 5438), adult male, collected at the administration buildings and campsite of Manombo Reserve, southeastern Madagascar, 23° 01.699' S, 47° 43.892' E, at 44 m above sea level, by M. Vences, G. Safarek, E. Rajeriarison and T. Rajoafiarison on 23 February 2007. ZMA 20097 ( ZCMV 660), ZMA 20129 ( ZCMV 662), UADBA uncatalogued ( ZCMV 661), three adult males from Vondrozo village (not morphologically examined for this study), 22° 49.183' S, 47°19.216' E, 198 m above sea level, collected by M. Vences and E. Rajeriarison on 9 February 2004.
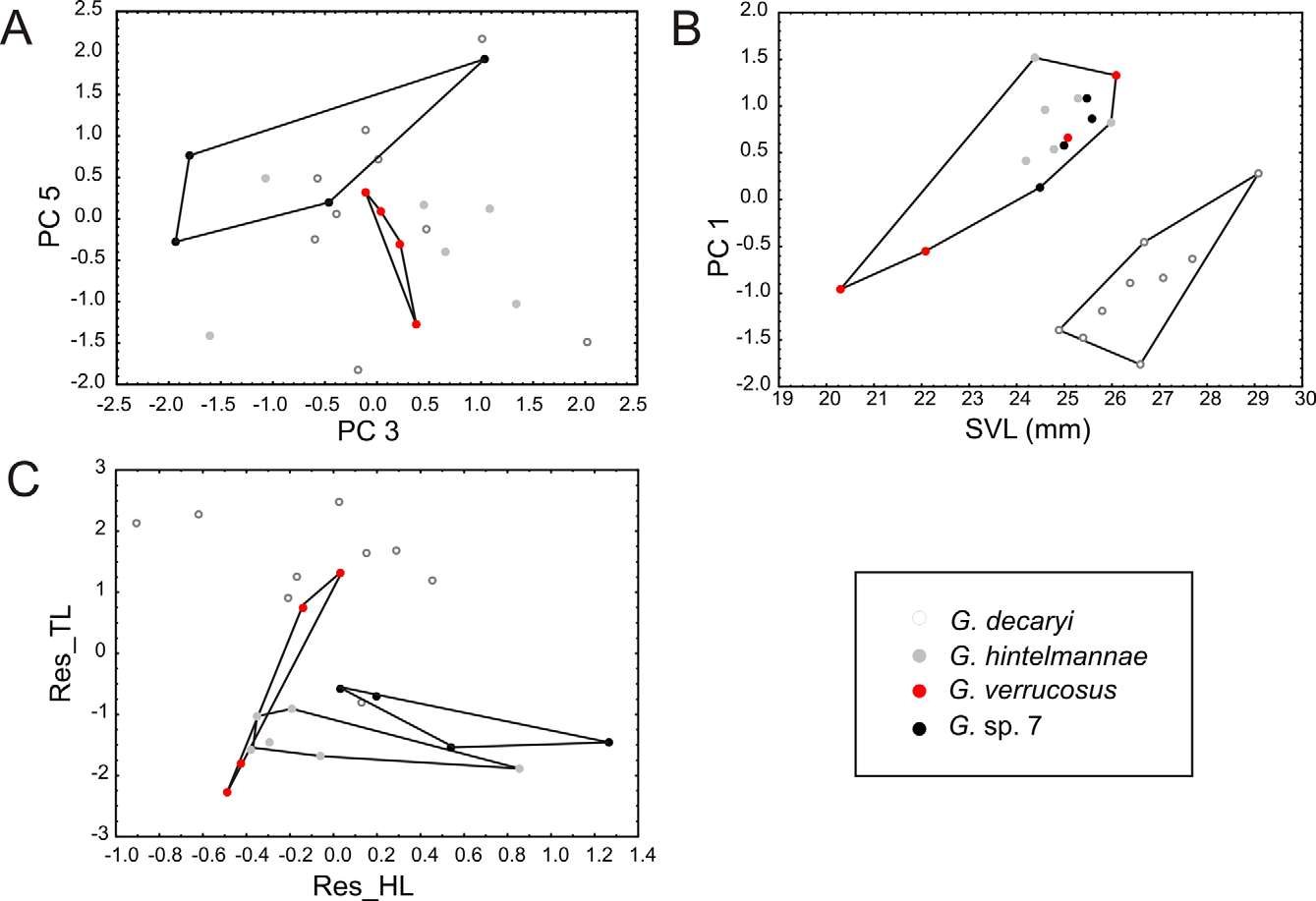
Diagnosis. A member of the subgenus Gephyromantis in the genus Gephyromantis on the basis of (1) presence of intercalary elements between ultimate and penultimate phalanges of fingers and toes (verified by external examination), (2) small size (SVL below 35 mm), (3) slightly enlarged terminal discs of fingers, (4) presence of outer metatarsal tubercle, (5) absence of webbing on hands and presence of only rudimentary webbing on feet, (6) tight connection of tissue surrounding the two lateral metatarsalia, (7) presence of femoral glands in males, (8) presence of paired or bilobed blackish vocal sacs in males, (9) diurnal calling behavior not concentrated at water bodies. Within the subgenus, distinguished from all other species by combination of (1) comparatively small to moderate size (male SVL 20–26 mm), (2) dorsum coarsely tubercular (the tubercles sometimes arranged such as to roughly form interrupted inner and outer dorsolateral ridges), (3) upper lip with light gray or white color interrupted by 2–4 distinct dark markings, (4) lower lip ventrally without a distinct alternating series of brown and yellowish markings; (5) dorsal color light gray with a contrasted, rather symmetrical dark pattern, (6) hindlimbs not particularly long, not reaching beyond snout tip, (7) calls consisting of a rather long series of>30 notes of indistinctly pulsed structure with a low note repetition rate of 3.6–4.6/s.
Assigned to the group of species here informally defined as the Gephyromantis decaryi complex based on its molecular phylogenetic relationships (see above) and its calls consisting of long series of notes with a low note repetition rate. Distinguished from G. decaryi by lacking inner and outer dorsolateral ridges (sometimes interrupted rows of tubercles present vs. distinct and usually not interrupted), by the small and externally indistinct femoral glands in males (vs. larger and more distinct), by shorter limbs (tibiotarsal articulation at most reaching snout tip vs. reaching beyond snout tip), and by a longer note duration (123–157 vs. 65–120 ms; see Table 2 View TABLE 2 ) in advertisement calls. Distinguished from G. hintelmannae (described below) by a much more granular dorsum (large regular granules versus smooth with small tubercles and traces of dorsolateral ridges, Fig. 6 View FIGURE 6 ), by the small and externally indistinct femoral glands in males (versus larger and more distinct), by the presence of distinct dark patches interrupting light color on upper lip (versus usually absence), by usually a contrasted dorsal pattern (versus usually uniform grayish), possibly by a lower number of notes per call (<80 versus>100), and by a longer note duration (123–157 vs. 81–103 ms; see Table 2 View TABLE 2 ) in advertisement calls. G. verrucosus is also well separated from all species of the G. decaryi complex, except G. hintelmannae , by morphometry ( Fig. 5 View FIGURE 5 ).
Redescription. Based on adult male specimen ZSM 2508/2007 from Manombo. Specimen in excellent state of preservation. A tissue sample has been taken for DNA analysis on the right femur. SVL 22.1 mm. For measurements see Table 1. Body slender, ratio of head length to head width HL/HW = 1.03, head slightly wider than body, snout pointed in dorsal view, rounded in lateral view; nostrils directed posterolaterally, slightly protuberant, canthus rostralis moderately distinct, rounded; loreal region concave; tympanum distinct, rounded, ratio of tympanum diameter to eye diameter TD/ED = 0.56; supratympanic fold prominent; tongue ovoid, posteriorly bifid; maxillary teeth present; vomerine teeth absent; choanae rounded. Arms slender, distinct single subarticular tubercles; inner and outer metacarpal tubercles distinct; fingers without webbing; relative length of fingers 1<2<4<3, fourth finger slightly longer than second finger; finger discs distinctly enlarged; nuptial pads absent. Hindlimbs slender; tibiotarsal articulation reaches the nostril when the hindlimb is adpressed along the body; lateral metatarsalia strongly connected; inner metatarsal tubercle distinct, outer metatarsal tubercle weakly recognizable; traces of webbing between toes; relative length of toes 1<2<5<3<4; fifth toe slightly shorter than third toe. Skin on the dorsal surface smooth, with numerous weakly developed, interrupted and inconsistently shaped dorsolateral and lateral ridges, scattered small tubercle-shaped ridges on the head ( Fig. 6 View FIGURE 6 ). Ventral skin smooth on throat, chest and limbs, granular on central, posterior and lateral belly. The subgular vocal sac is laterally black, probably indicating a paired or bilobed shape when inflated; femoral glands are very weakly developed, and indistinct in external view.
After almost three years in preservative, dorsally uniformly dark brown with diffuse dark markings on the dorsum. On the flanks, the dorsal color gradually fades into the light ventral coloration. The tympanic area is dark brown; the supratympanic fold is also dark. Under the eye, a whitish spot is bordered by two dark stripes. No white lip stripe is present. Dark-brown crossbands on the limbs. Ventrally, the color is light tan, shanks and thighs are brownish-mottled. A white irregular median band, bordered by tan coloration, runs along the throat and chest, which also has some additional white pigment. The lower lip shows ventrally a distinct alternating brown-white pattern (two white spots on each side). The anterior belly has some white mottling on tan background. Color in life ( Fig. 4 View FIGURE 4 g) very similar to that in preservative. Dorsally, the ground color was gray-brown, markings and pattern brown and dark brown. Ventrally, the light stripe on the throat and light marbling on chest were of a distinct silvery white. Some very weak yellowish tan was present on the posterior belly, in the ventral part of the inguinal region, and on the femoral glands. The iris was rather uniformly light brown, with a small dark brown spot centrally in its lower half.
Variation. For differences in body size measurements see Table 1. Specimens ZSM 2462/2007 (ZCMV 5459), ZSM 2506/2007 (ZCMV 5450) and ZSM 2507/2007 (ZCMV 5451) have been dissected ventrally (presumably to check for internal parasites). Ventral coloration of ZSM 2505/2007 (ZCMV 5438) darker than in holotype; ZSM 2507/2007 (ZCMV 5451) showing a highly contrasted dorsal pattern (but structurally similar to holotype); ZSM 2505/2007 (ZCMV 5438) having more strongly developed, smaller tubercle-like interrupted dorsolateral and lateral ridges and also above the eyes; color of ZSM 2505/2007 (ZCMV 5438) in preservative is a more reddish brown than in all other specimens.
Natural history. In the village of Vondrozo, specimens were calling during the day in gardens and among shrubs, in a completely deforested area with single large fruit trees. At Manombo Special Reserve, specimens were calling in the afternoon from shrubs and logs at low perch heights (<50 cm) in the forest, and in dense fern vegetation at the forest edge. Calls were also emitted during dusk and sporadically also heard after nightfall. At Manombo, calling specimens clearly were more common in clearings or at the forest edge than in dense forest. Calling males were evenly spaced. Reproduction and larval stages are unknown but the fact that calling males are not concentrated close to water bodies suggests a possible nidicolous reproduction as in other species of the subgenus Gephyromantis .
Advertisement calls. Two calls from Vondrozo consisted of 31 and 40 notes each. Intensity of notes increases from the first notes on, and slightly decreases again towards the end of the call. Note repetition rate was 3.7/s. Notes had the most clearly pulsed structure of all populations in the G. decaryi complex studied here, with 24–29 pulses/note (n=3). A fundamental frequency band is observed between 1800–2800 Hz, whereas dominant frequency ranges between 3800–5300 Hz.
The two analyzed calls from Manombo consist of 50, 64 and 71 notes arranged in a regular series plus three notes emitted at somewhat longer intervals at the end of a call. Note repetition rate was 4.5/s. Notes were less distinctly pulsed than at Vondrozo, with 30–35 recognizable pulses (n=3).
Distribution. Known from three low and mid-altitude localities in south-eastern Madagascar: (1) the type locality Fort Carnot ( Ikongo) at 600 m a.s.l., (2) Manombo Special Reserve (ca. 50 m a.s.l.), and (3) the village Vondrozo (ca. 200 m a.s.l.). We also heard calls probably referable to this species from along the road running from Vondrozo to the nearby Vevembe forest.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |