
Furculanurida emucronata, Zon, Serge Demeango, Tano, Yao & Deharveng, Louis, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3878.5.8 |
|
publication LSID |
lsid:zoobank.org:pub:A6A3020B-75AC-434C-83DF-60FCD6171680 |
|
DOI |
https://doi.org/10.5281/zenodo.5698265 |
|
persistent identifier |
https://treatment.plazi.org/id/03C66869-FFE0-341E-88C8-35384878B281 |
|
treatment provided by |
Plazi |
|
scientific name |
Furculanurida emucronata |
| status |
sp. nov. |
Furculanurida emucronata sp. nov.
( Figs 1–15 View FIGURES 1 – 2 View FIGURES 3 – 9 View FIGURE 10 View FIGURES 11 – 15 )
Type material. Holotype female and one paratype male on slides: Ivory Coast, near Lamto Reserve, 10 July 2013, litter in Theobroma cacao L. (cocoa) plantation, S. Zon leg., 06°15’43.6”N, 05°01’29.8”W, elevation 145 m, Berlese/Tullgren extraction (sample VC01L8). One paratype male: same data, except collected in a pitfall trap (sample VC01P4). One paratype female juvenile: same locality, 26 October 2012, soil from a three-four years fallow, 06°15’39.9”N, 05°01’29.9”W, elevation 139 m (sample VJ31S3).
Holotype and one paratype in Laboratoire de Zoologie et Biologie Animale de l’Université Félix Houphouët- Boigny (Abidjan, Ivory Coast); two paratypes in Museum National d'Histoire Naturelle (Paris, France).
Description. Length: 0.80 to 1.30 mm (holotype: 1.30 mm). Colour white in ethanol. Habitus stocky, paratergites not developed, antennae slightly shorter than head ( Figs 1–3 View FIGURES 1 – 2 View FIGURES 3 – 9 ). Secondary granulation coarse and regular. No tubercles. No reticulations. No plurichaetosis. Chaetotaxy symmetrical with rare exceptions ( Figs 3 View FIGURES 3 – 9 , 13 View FIGURES 11 – 15 ).
Mouthparts ( Figs 6–7 View FIGURES 3 – 9 ). Maxilla styliform, thin, minutely hooked apically; mandibular head swollen before apex, slightly bent distally, with one row of unequal teeth arranged as follows from base to apex: 2 large, 1 small, one large, 2 small, 1 large ( Fig. 6 View FIGURES 3 – 9 ). Labium with 4 basal chaetae, 4 distal chaetae and at least 2 lateral chaetae, distally armed with a pair of strong symmetrical internal bidentate spines in both sexes, labrum short, broad, truncated apically ( Fig. 7 View FIGURES 3 – 9 ); labral formula 4/3,5,4.
Antennae ( Figs 8, 9 View FIGURES 3 – 9 ). First antennal segment with 7 chaetae, second segment with 11 chaetae. Third and fourth antennal segments fused dorsally. Sensory organ of third antennal segment consisting of two short swollen, smooth S-chaetae; two long, subcylindrical S-chaetae Sgv and Sgd; and one S-microchaeta (ms) ventrally; 18 ordinary chaetae on the segment, acuminate, most located ventrally. Fourth antennal segment dorsally with a trilobed apical bulb, a short chaeta “i”, 7 curved S-chaetae of same length as guard S-chaetae of Ant. III, several macrochaetae of “mou” type and a small, broad, rounded subapical organite (or) ending in several micro-teeth; dorso-lateral Smicrochaeta (ms) absent. No ventral sensory rasp on Ant. IV.
Ocular plate. Eyes absent. PAO elliptical, constituted of 13–16 vesicles arranged in a single row; each vesicle oval-elongate, variously lobed, giving to the same PAO a moruliform or normal (non-moruliform) aspect when examined at different focus level ( Fig. 10 View FIGURE 10 ).
Head and body chaetotaxy ( Figs 3, 4–5 View FIGURES 3 – 9 , 13, 15 View FIGURES 11 – 15 ; Tab. 1 View TABLE 1 ). Dorsal ordinary chaetae short, pointed, subequal; no macrochaetae; S chaetae of tergites rather thin and long, more than six times longer than closest ordinary chaetae, arranged as 2+ms,2/1,1,1,1,1 by half-tergite from Th. II to Abd. V ( Fig. 3 View FIGURES 3 – 9 ). On Th. II, dorso-lateral ms rod-shape, located internally to S ( Fig. 4 View FIGURES 3 – 9 ). Presence of 1+1 pouch-like structures posteriorly on head and dorso-laterally on Abd. I ( Fig. 5 View FIGURES 3 – 9 ), similar to those observed in some Neanuridae ( Deharveng 1983b) . Chaetal groups clearly recognisable on tergites from Th. I to Abd. III ( Fig. 3 View FIGURES 3 – 9 ; Tab. 1 View TABLE 1 ).
On Th. II and Th. III, the 3 chaetae Di on a same longitudinal line; Abd. V with 2+2 chaetae (a1, p1) between S-chaetae ( Fig. 3 View FIGURES 3 – 9 ). Head with five post-labial chaetae, including 2 per side along linea ventralis. No thoracic chaetae ventrally. Chaetotaxy of abdominal sternites illustrated in Fig. 13 View FIGURES 11 – 15 . Each valve of Abd. VI with 3 mic, the median one often difficult to detect. Male genital plate with 15–20 small smooth acuminate circumgenital mesochaetae around the periphery and 4+4 central smooth pointed genital microchaetae ( Fig. 15 View FIGURES 11 – 15 ).
Body appendages ( Figs 11, 12, 14 View FIGURES 11 – 15 ). All tibiotarsal chaetae straight or slightly curved, acuminate; tibiotarsi I, II, III with 19, 19, 18 chaetae (M present, displaced basally); ventral macrochaetae not overpassing the apex of tibiotarsus ( Fig. 12 View FIGURES 11 – 15 ). Trochanter with 6 chaetae each, including a thinner one. Claw of leg I with a very strong lateral tooth inserted basally; claws of leg II and of leg III with a short thin pointed inner tooth inserted at 1/2–1/3 of the claw, devoid of lateral tooth ( Fig. 11 View FIGURES 11 – 15 ). Ventral tube with 3 chaetae on each flap. Rami of tenaculum with 3 teeth each, the distal 1+1 smaller. Furca reduced, dens globular with 6 chaetae, mucro absent ( Fig. 14 View FIGURES 11 – 15 ).
Name derivation. The species name refers to the absence of mucro. Ecology. The species was rare in our samples, and restricted to cocoa plantations and fallow ground. It was absent in forest and savannah.
Discussion. Furculanurida emucronata sp. nov. is well characterised by several morphological features. For instance, the unusually strong lateral tooth on the claw of anterior pair of legs is unique for species of Pseudachorutinae . The absence of mucro is unique among Furculanurida , and the absence of eyes is only shared with F. a f r i c a na Massoud, 1963 in the genus. On the other hand, the morphology of the new species suggests relationships with several other genera of Pseudachorutinae , raising questions about the current generic structure of the subfamily.
Furculanurida emucronata sp. nov. has been provisionally assigned here to the genus Furculanurida , on the ground of its similarity with F. africana , the type species of the genus, from which it differs only by fewer vesicles in PAO and the absence of mucro. However, this assignment raises two problems. Firstly, the genus Furculanurida , already poorly defined and overlapping in morphology with Pseudachorutes (because of F. tropicalia Queiroz & Fernandes, 2011 ), would now overlap also with Stachorutes (because of F. emucronata sp. nov.). Secondly, the new species exhibits several characteristics that indicate close relationships with the genera Arlesiella and possibly also to the African genus, Kenyura.
The genus Furculanurida was established by Massoud (1967: 144) within Pseudachorutinae for the species Micranurida africana from Ivory Coast, on the basis of the following combination of characters: PAO present, maxilla styliform, furca present and reduced number of eyes. It was re-diagnosed by Thibaud & Palacios-Vargas (2000) and Queiroz & Fernandes (2011). These last authors include 13 nominal species in the genus, distributed in the tropics of Africa and America, with a single Nearctic record. In the same paper, they described F. tropicalia , a species with 8+8 eyes that complies with the genus Pseudachorutes Tullberg, 1871 rather than Furculanurida according to the definition of these genera at that time. However, in several other characters, F. tropicalia was considered closer to South American Furculanurida than to Pseudachorute s and so was assigned to the former genus. In support to this view, the authors compared Pseudachorutes and Furculanurida in detail and concluded that they only differed in habitus. However, the presence/absence of ms on Ant. IV, not known for several species in either genus, including the generotype of Furculanurida , may be a better diagnostic character.
Queiroz & Fernandes (2011) wrote “The habitus of Furculanurida differs markedly from the habitus of Pseudachorutes and Stachorutes , which are typically of type 1 (after Massoud, 1967: 144). The Furculanurida habitus is of type 2, i.e. a stout and flattened body with Abd V tending to be deformed and Abd VI visible dorsally”. Type 1 habitus is characterised by a complete absence of paratergites. Type 2 habitus differs from type 1 by the presence of weakly developed paratergites, while type 3 and 4 have well developed paratergites. These categories are not always easy to recognize as stated by Massoud himself (1967: 187), who did not give them discriminant value at generic level among Pseudachorutinae . Regarding Furculanurida, Massoud (1967) mentions in its diagnosis “sans paratergite saillant”. In the description of Micranurida africana , the type species of the genus, Massoud (1963) does not mention the presence of paratergites, and the habitus he figures is very similar to that of a Pseudachorutes , i.e. without paratergites. As a result, the presence/absence of paratergites alone does not allow separation of Furculanurida from Pseudachorutes .
Pseudachorutes species with reduced furca and reduced number of eyes have been separated into the genus Stachorutes by Dallai (1973). Comparing Stachorutes and Furculanurida, Thibaud & Palacios-Vargas (2000) , suggested that they could be differentiated by the number of mandibular teeth, number of S-chaetae on Ant. IV, length of S-chaetae on tergites, presence/absence of inner tooth on claw and furcal development although these authors do not formally list differences. The number of S-chaetae on Ant. IV in Furculanurida species, when known, is 6 or 7, not 8 as stated by the authors, versus 5 or 6 in Stachorutes ; hence it is not a diagnostic character. Inner tooth on claw and number of mandibular teeth are usually considered as specific, not generic characters. Furcal development is not a generic character in Neanuridae according to Massoud (1963) at least where dentes are developed; large genera with species at different degree of furcal reduction, like Aethiopella or Friesea , are not exceptional. The length of S-chaetae on tergites allows Furculanurida of tropical Africa (with very long S-chaetae, at least 5 times longer than closest ordinary chaetae) to be discriminated from Stachorutes species (shorter Schaetae), but is not relevant for several non-African Furculanurida . As the presence of paratergites cannot be considered as a diagnostic morphological trait of Furculanurida , there is no differential character available to distinguish this genus and Stachorutes .
The genus Furculanurida can only be separated from Pseudachorutes by a ‘conditional’ combination of characters, i.e. “eyes fewer than 8+8, or, when 8+8, microchaeta ms absent on Ant. IV”. Similarly, difference with Stachorutes would be: “mucrodens complete, or, if mucro absent, ms absent on Ant. IV”.
A second taxonomical problem is the close relationships between F. emucronata sp. nov. and species of the genera Arlesiella and Kenyura. The new species differs unambiguously of these two genera by the presence of a PAO, but it also shares with them original characters of mouthparts, antennae and ventral tube. The head of mandible is externally swollen in F. emucronata sp. nov., F. furculata ( Salmon, 1956) and F. grandcolasorum Weiner & Najt, 1998 , but also in all species of Arlesiella and in African species of Kenyura. The presence of spines distally on labium, is only known so far in Arlesiella and F. emucronata sp. nov. However, this character may have been overlooked in species description of Furculanurida and Kenyura, which do not include details of the labial distal edge. Where known, the absence of ms on Ant. IV ( Queiroz & Fernandes 2011) and the presence of only 3+3 chaetae on ventral tube are also shared between Arlesiella and African species of Furculanurida , although both characters remain unknown in the type species of the latter genus. A redescription of this type species is needed to redefine the genus Furculanurida and to check its relationships with Arlesiella and Kenyura. The single type specimen of F. africana ( Massoud, 1963) preserved in the MNHN, is in a poor condition, with only mandibles clearly observable. Recent sampling in the Banco forest did not provide any F. africana so without fresh topotypic specimens, the assignment of the species emucronata sp. nov. to Furculanurida has to be considered provisional. The same holds for all species of the genus with the exception of the type species.


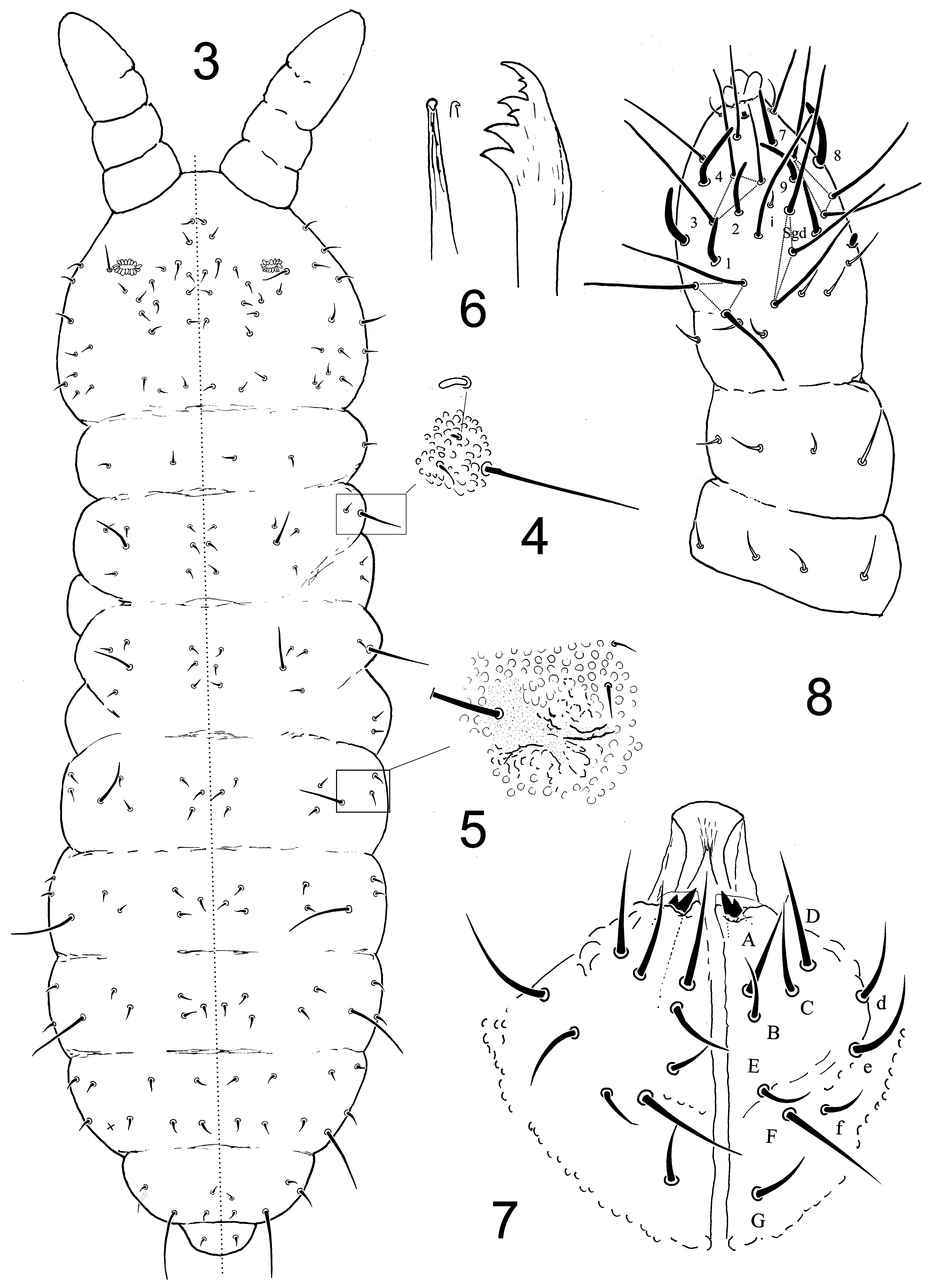

TABLE 1. Chaetotaxy of tergites from Th. I to Abd. III, by chaetal groups of Furculanurida emucronata sp. nov.
| Di | De | DL | L | |
|---|---|---|---|---|
| Th. I | 1 | 1 | ||
| Th. II | 3 | 3+S | 1+S+ms | 2 |
| Th. III | 3 | 3+S | 1+S | 2 |
| Abd. I | 3 | 2+S | 2 | 2 |
| Abd. II | 3 | 2+S | 2 | 2 |
| Abd. III | 3 | 2+S | 2 | 2 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Furculanurida emucronata
| Zon, Serge Demeango, Tano, Yao & Deharveng, Louis 2014 |
F. tropicalia
| Queiroz & Fernandes 2011 |
Furculanurida
| Thibaud & Palacios-Vargas 2000 |
F. grandcolasorum
| Weiner & Najt 1998 |
Furculanurida
| Massoud 1967 |
F. africana (
| Massoud 1963 |
F. furculata (
| Salmon 1956 |
Pseudachorutes
| Tullberg 1871 |