
Eurypon suassunai, Santos, George Garcia, França, Fernando & Pinheiro, Ulisses, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3895.2.8 |
|
publication LSID |
lsid:zoobank.org:pub:12702F5B-4E55-4CCA-8664-1F6269EF06EA |
|
DOI |
https://doi.org/10.5281/zenodo.6123199 |
|
persistent identifier |
https://treatment.plazi.org/id/A108FC07-FFEB-FFB5-FDF5-FE3FC44DF929 |
|
treatment provided by |
Plazi |
|
scientific name |
Eurypon suassunai |
| status |
sp. nov. |
Eurypon suassunai sp. nov.
( Figs 1 View FIGURE 1 , 4–5 View FIGURE 4 View FIGURE 5 ; Table 1)
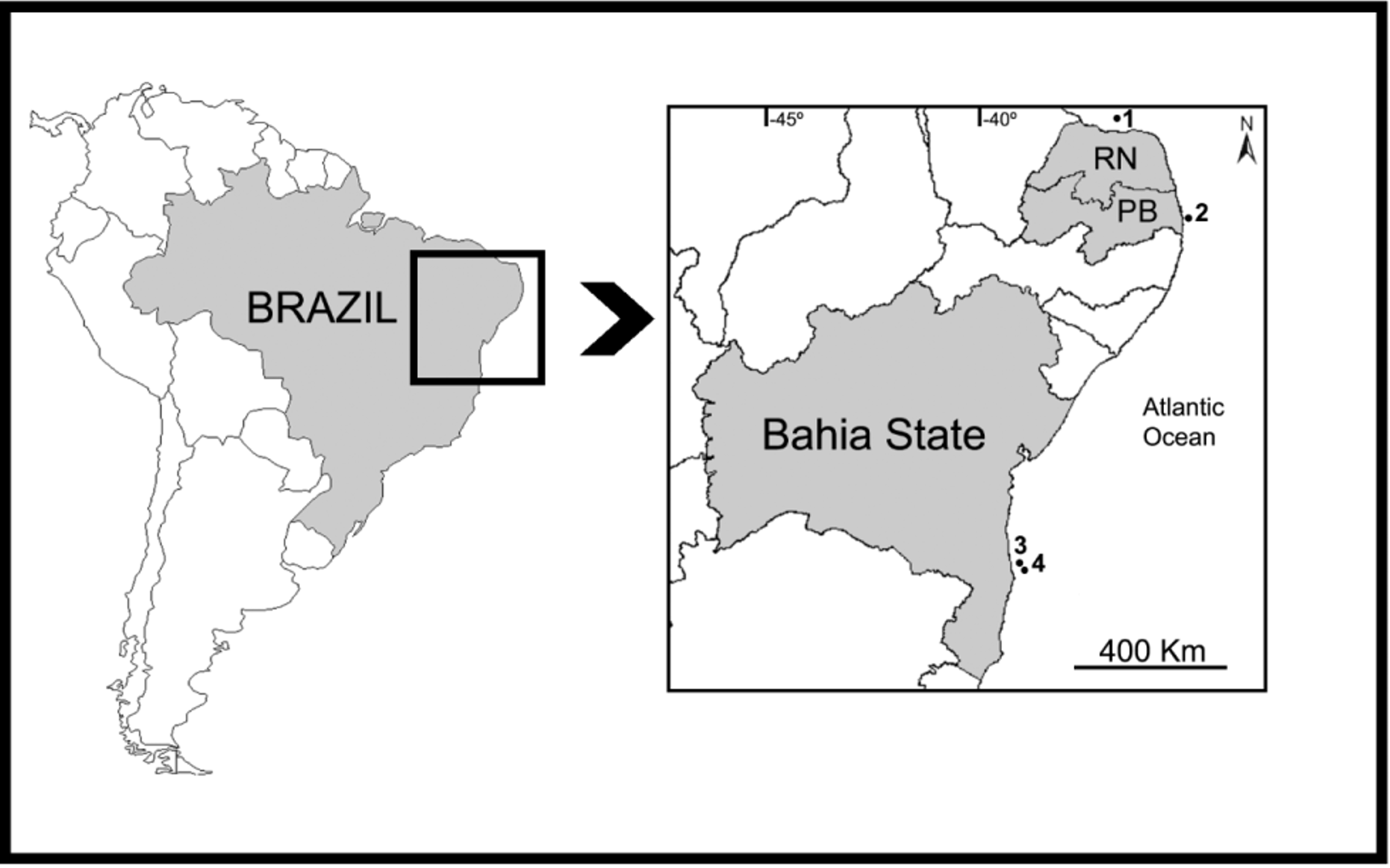
Type specimens: UFPEPOR 1532 (Holotype), off Canavieiras City, Camamu-Almada Basin (15° 35' 5.91" S, 38° 46' 36.35" W), Bahia State, Brazil, (X.2011). UFPEPOR 1533 (paratype), off Canavieiras City, Camamu-Almada Basin (15° 33' 30.56" S, 38° 45' 14.53" W), Bahia State, Brazil, (X.2011).
Diagnosis. Eurypon suassunai sp. nov. is the only Eurypon from the Atlantic which combines subtylostyles, acanthostyles I, acanthostyles II with bulbous base, and raphidiform styles.
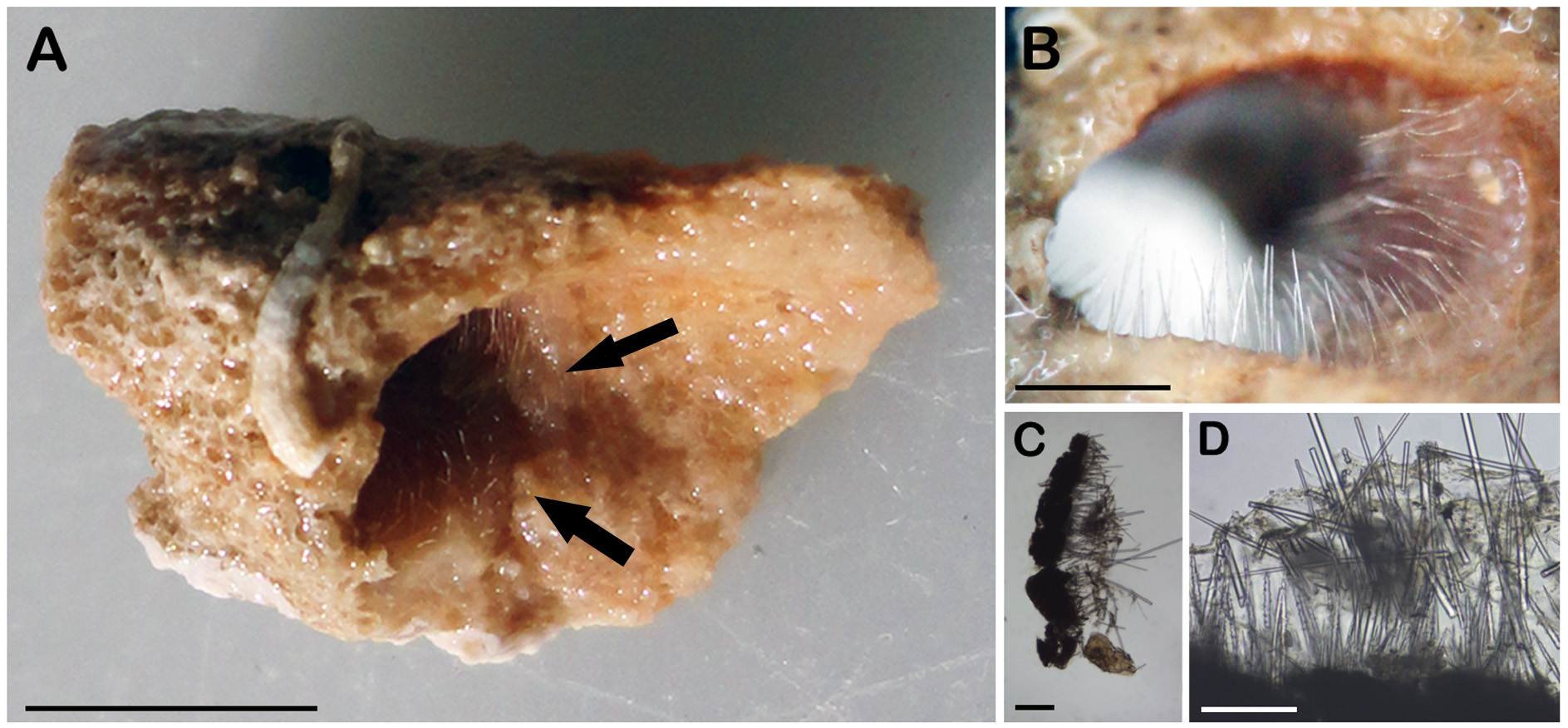
External morphology ( Fig 4 View FIGURE 4 A–B). Thin transparent encrustation, about 1 mm thick. Oscules not visible. Surface hispid due to evenly distributed projecting spicules. Consistency is fragile. Color in life is orange (formaldehyde) and pale in ethanol.
Skeleton ( Fig. 4 View FIGURE 4 C–D). Ectosomal skeleton is absent. Tips of the choanosomal spicules protrude externally ( Fig. 4 View FIGURE 4 C–D). The choanosomal skeleton has a hymedesmioid structure. The raphidiform styles are dispersed in the subectosomal region. Acanthostyles I and II are most abundant on basal layer of spongin ( Fig. 2 View FIGURE 2 D).
Spicules ( Fig. 5 View FIGURE 5 A–I). Choanosomal subtylostyles ( Fig. 5 View FIGURE 5 A, G): long, smooth, slender, slightly curved and with bulbous base (690–1660 / 5–17 µm); Acanthostyles I ( Fig. 5 View FIGURE 5 B, E): varying from straight to slightly curved, with lightly bulbous base, short and curved spines (hook-shaped spines) more concentrated in the middle and apical part of spicules (159–354 / 7–13 µm); Acanthostyles II ( Fig. 5 View FIGURE 5 C, F): are short and robust, often straight, with bulbous base, totally microspined with short and curved spines (hook-shaped spines) (54–129 µm); Raphidiform styles ( Fig. 5 View FIGURE 5 D, H, I): smooth, thin, most are straight, the styloid base ranges from the shape of a crown (171–345 / 1–5 µm).
Distribution ( Fig. 1 View FIGURE 1 ). Northeastern coast of Brazil, Bahia State, Brazil.
Depth. Shallow water.
Etymology. This species is named in honor of the Late the famous writer Ariano Vilar Suassuna for his defense of the culture of the Brazilian Northeast.
TABLE 1. Comparative micrometric data on the spicules and overview of distribution of the living species of Eurypon Gray, 1867 for the Atlantic. Values are in micrometers (µm), expressed minimum–maximum or minimum– mean –maximum length/width. References are numbered in parentheses and listed at the foot of the table.
TABLE 1. (continued)
Species Specimen Depth (m) Spicules (µm) location
Acanthostyles Subectosomal Choanosomal Choanosomal Other spicules Styles Tylostyles Subtylostyles
E. lictor ( Topsent, 1904) (3) Azores 1600 - - 2000 / 23 - 575–1000 / 4–5 (tornote) 50 / 13–15 (trichodragmata)
E. mixtum ( Topsent, 1928) (5) Ceuta, Spanish 650– 2165 I) 1000 - Not recorded Not recorded - Morocco II) 100–320
E. major Sarà & Siribelli, 1960 (10) Naples and 14–40 80–220 / 4–10.5 - 1115–2210 / - 480–700 / 4–7.5 (oxea)
Western Europe 10–17
E. mucronale ( Topsent, 1928) (5) Azores 2460 I) 700–900 / 25– - - Not recorded 400–490 / 12–17 (tornote)
30
II) 125–280 / 20
E. obtusum Vacelet, 1969 (10) Mediterranean 250 70–170 / 5–7.5 - Not recorded / - 400–430 / 2.5–3 (oxea)
Sea 10–12
E. pilosella ( Topsent, 1904) (3) Azores 550– 1360 I) 2000–3000 / - - 300–480 / 5–6 - 25–30 (microspined) II) 170–300 / 15–
25
E. radiatum ( Bowerbank, 1866) (11) Azores, Shetlands Not recorded 100–400 I) 800 / 10–15 - - - and Hebrides II) 350
E. scabiosum ( Topsent, 1927) (6) Azores 650–919 I) 1000 / 24 - - 470–630/3– 4 - II) 80–300 / 6–13
E. simplex (Bowerbank, 1874) (7) Roscoff and 60–260 I) 218 / 8 - - 2116 / 27 - Faroes II) 105
E. topsenti ( Burton, 1954) (13) Western 900 - 1200 / 8 1600 / 14 - 60 (trichodragmata)
Caribbean
E. toureti ( Topsent, 1894) (8) Gulf of Mexico Not recorded 50–60 - Not recorded - 10–12 (isochelae)
E. viride ( Topsent, 1889) (9) Gulf of Mexico, 80–800 85 - 1000 - 70 (raphide)
Mediterranean
and Azores
References: (1) Little (1963); (2) Lévi (1969); (3) Topsent (1904); (4) Topsent (1891); (5) Topsent (1928); (6) Topsent (1927); (7) Bowerbank (1874); (8) Topsent (1894); (9) Topsent (1889); (10) Aguilar-Camacho & Carballo (2013); (11) Bowerbank (1866); (12) Arndt (1935); (13) Burton (1954).
Remarks. Eurypon suassunai sp. nov. belongs to the genus in having choanosomal subtylostyles, echinating acanthostyles and an encrusting habit with a hymedesmioid skeleton. The closest species to E. suassunai sp. nov. is E. clavilectuarium sp. nov. because both shares the same spicules. However, in E. clavilectuarium sp. nov. the subtylostyles are longer and stouter (1200–2000 / 8–24) than those of E. suassunai sp. nov. (690–1600 / 5–17) and in the former, the tyle of the acanthostyles II are styloid against bulbose tyle of the last. Except from both, no others species of Eurypon in Atlantic have raphidiform styles. Furthermore, the presence of two categories of acanthostyles distinguishes E. suassunai sp. nov. from eleven species of the Atlantic: E. cinctum ; E. clavatella ; E. coronula ; E. fulvum ; E. lacazei ; E. lictor ; E. major ; E. obtusum ; E. radiatum ; E. toureti ; E. viride (Table 1).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Eurypon suassunai
| Santos, George Garcia, França, Fernando & Pinheiro, Ulisses 2014 |
E. obtusum
| Vacelet 1969 |
E. major Sarà & Siribelli, 1960
| Sara & Siribelli 1960 |
E. topsenti (
| Burton 1954 |
E. mixtum (
| Topsent 1928 |
E. mucronale (
| Topsent 1928 |
E. scabiosum (
| Topsent 1927 |
E. lictor (
| Topsent 1904 |
E. pilosella (
| Topsent 1904 |
E. toureti (
| Topsent 1894 |
E. viride (
| Topsent 1889 |
E. simplex
| Bowerbank 1874 |
E. radiatum (
| Bowerbank 1866 |