
Eurynome longimana Stimpson, 1857
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5048.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:951BE302-C0BF-4AA3-AE12-BBAC4EDEBAFB |
|
DOI |
https://doi.org/10.5281/zenodo.5569404 |
|
persistent identifier |
https://treatment.plazi.org/id/03B187B7-FF9E-FFB5-89B3-A5F7FAEB5C94 |
|
treatment provided by |
Plazi |
|
scientific name |
Eurynome longimana Stimpson, 1857 |
| status |
|
Eurynome longimana Stimpson, 1857 View in CoL
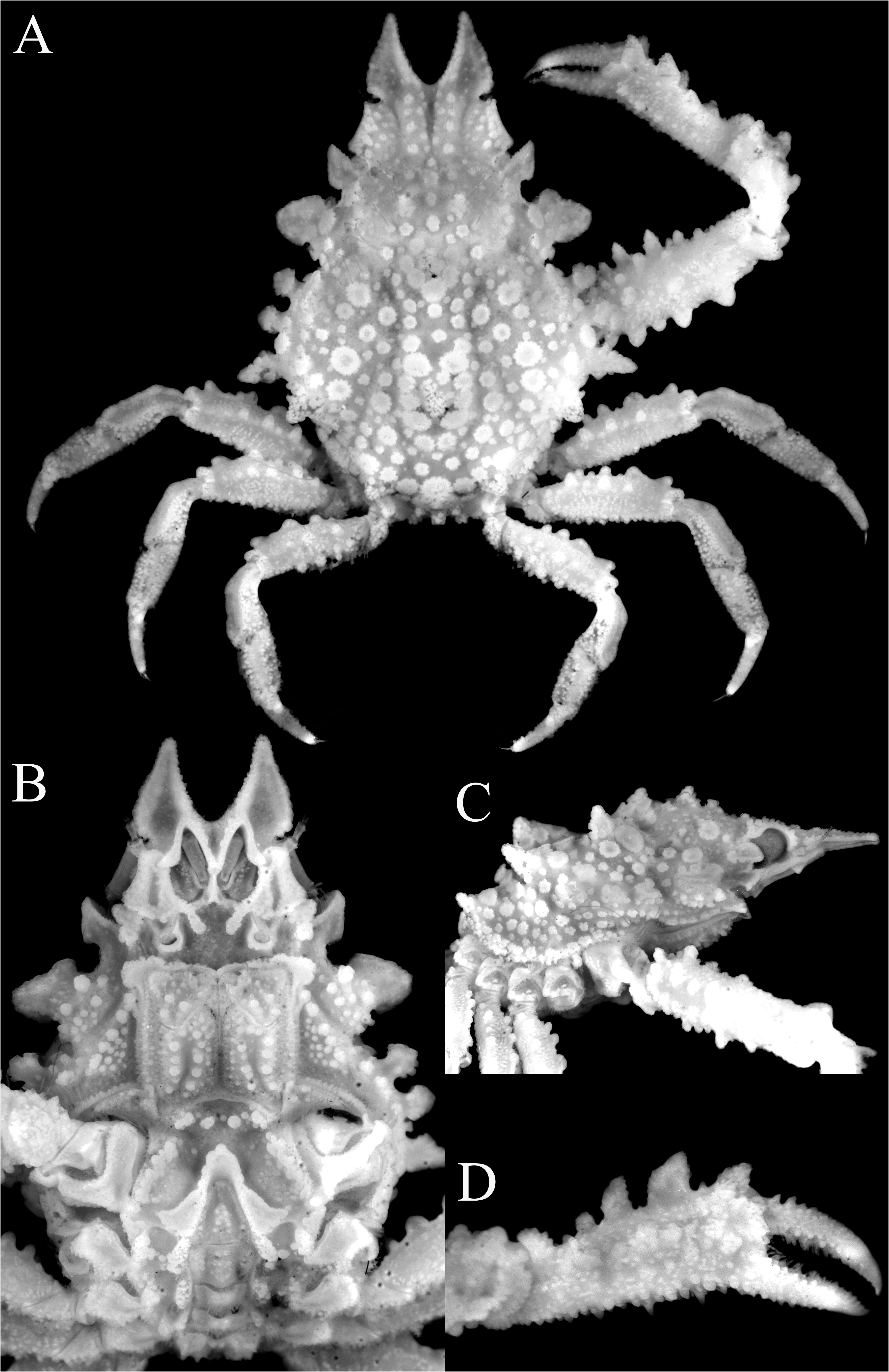
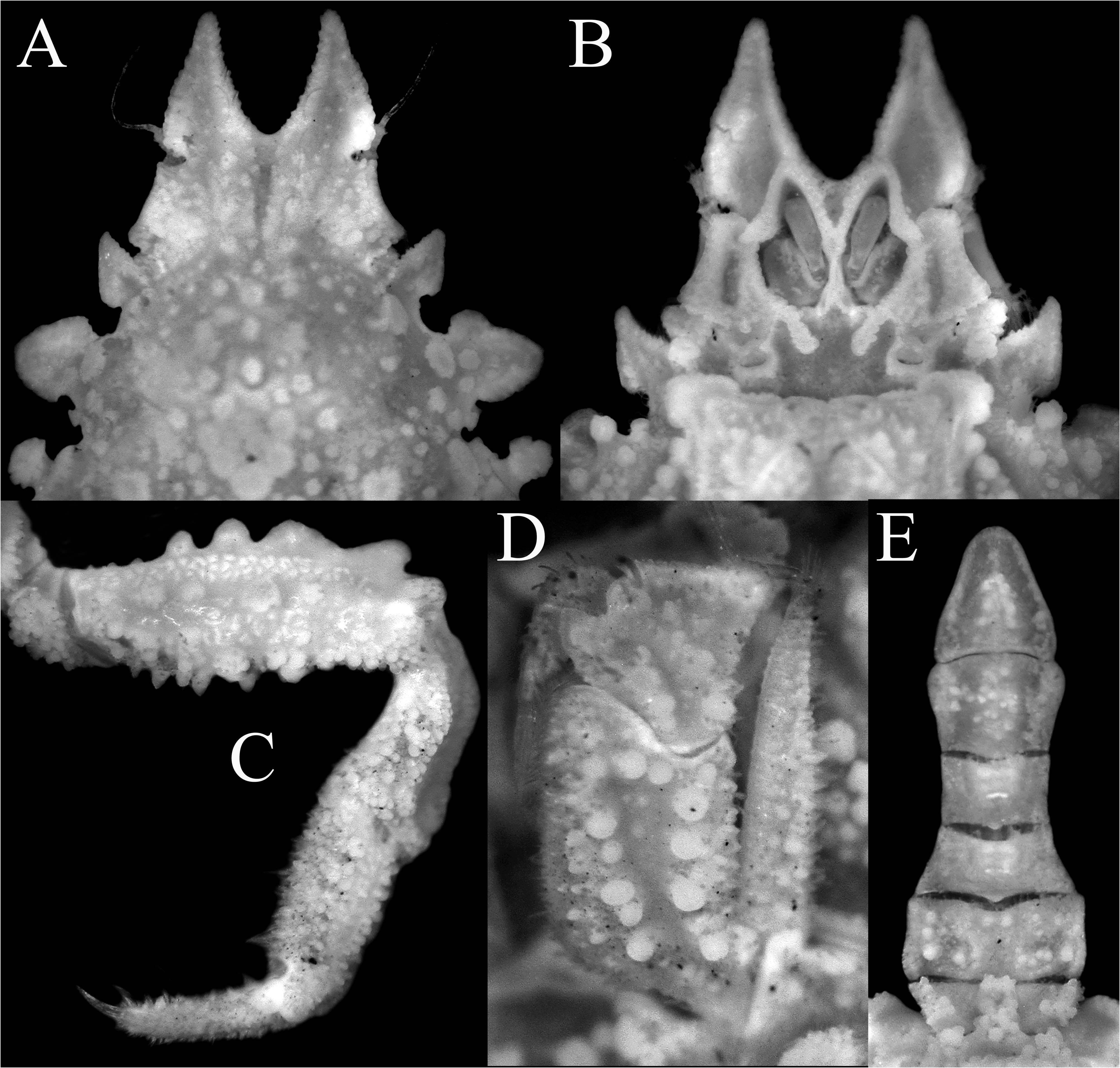
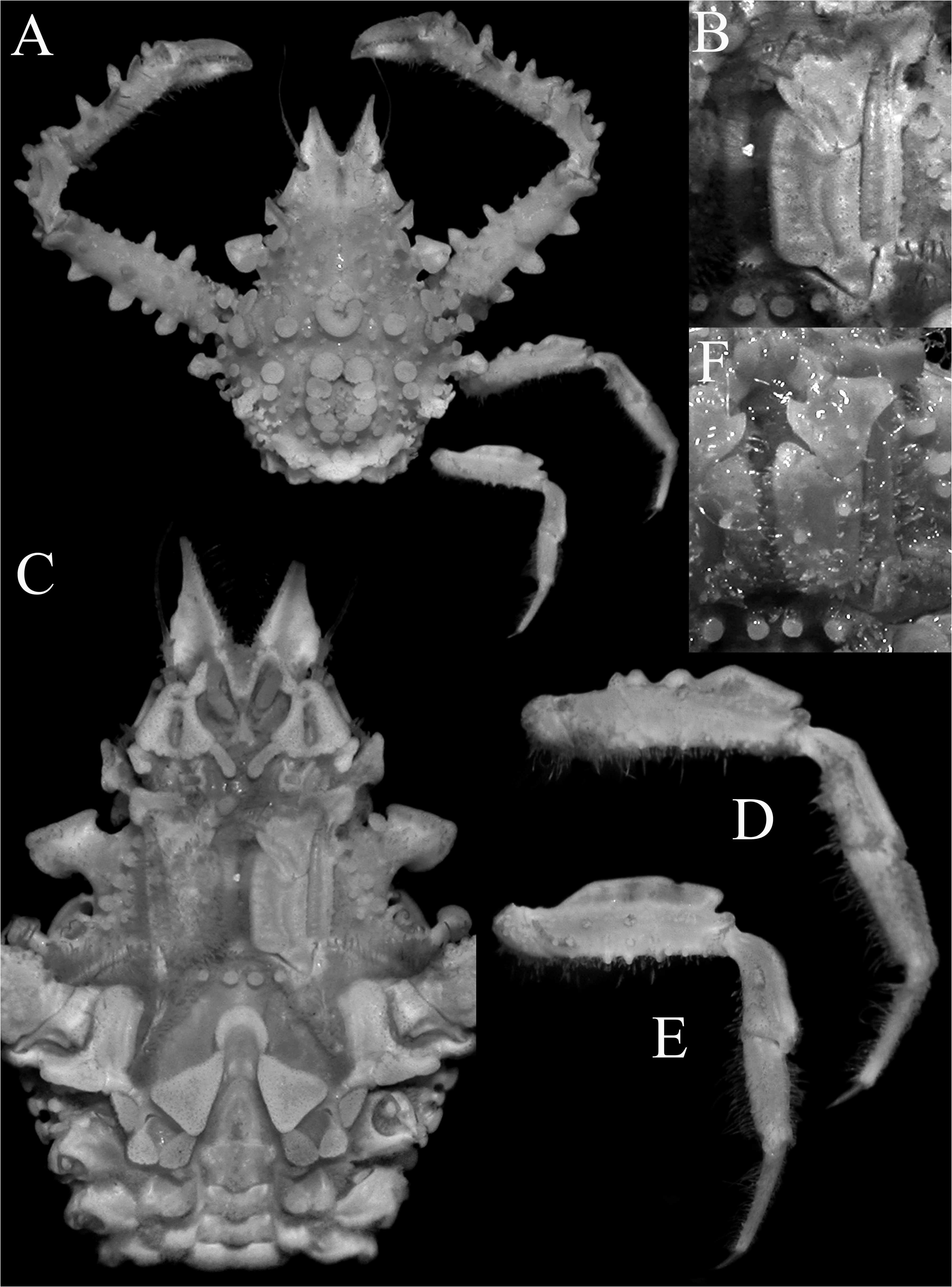
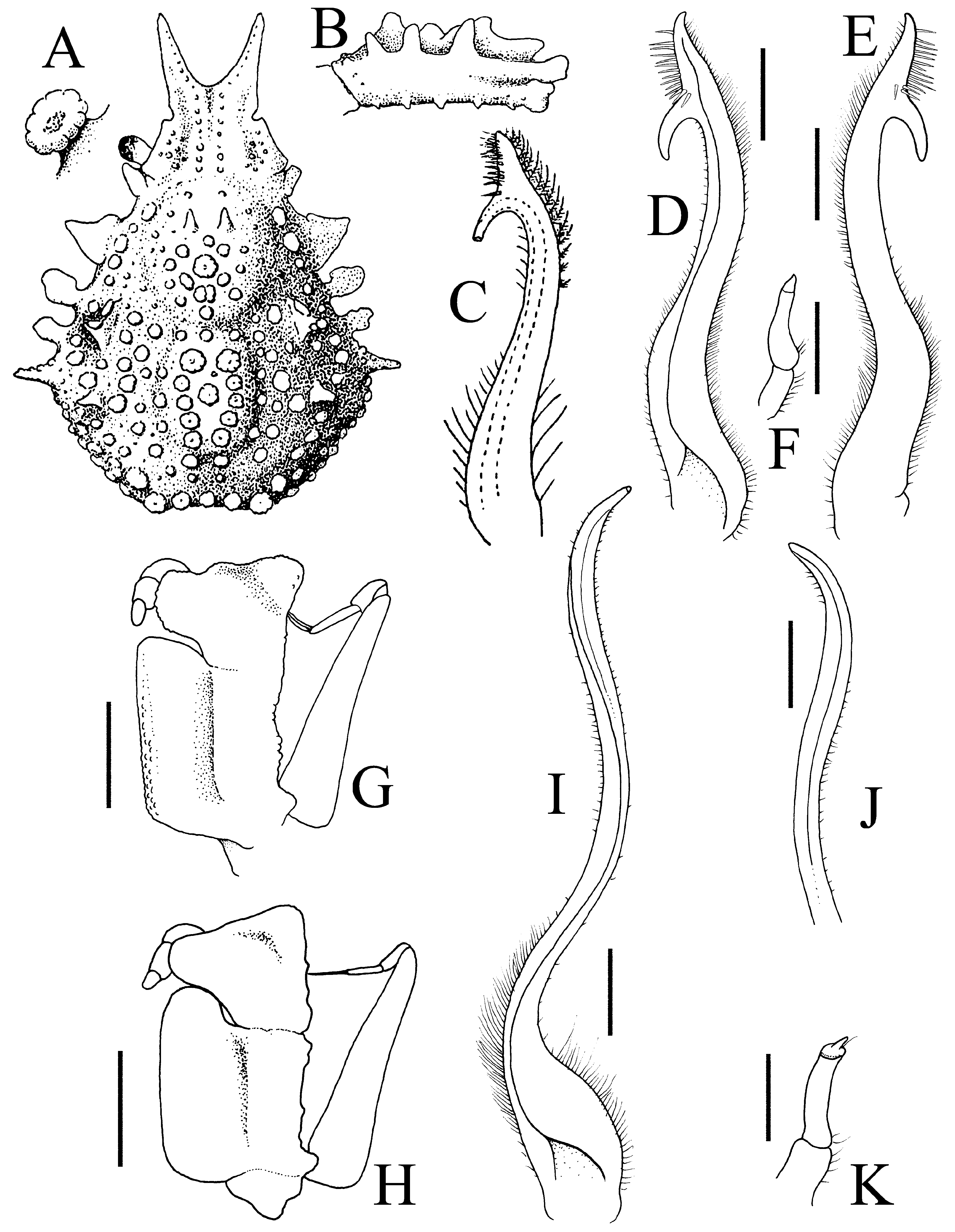
( Figures 2 View FIGURE 2 , 3 View FIGURE 3 , 13A–F View FIGURE 13 )
Eurynome longimana Stimpson, 1857: 220 View in CoL .— Rathbun, 1893: 102, pl. 8 fig. 1.— Stimpson, 1907: 27, pl. 4 fig. 2.— Stebbing, 1910: 289.— Lenz, in Lenz & Strunck, 1914: 275.— Capart, 1951: 87; Monod, 1956: 482.
Eurynome aspera View in CoL — Barnard, 1950: 56; fig. 12a–c (not Cancer asper Pennant, 1777 ).
Material examined. 1 male (cl 8.5 mm, pcl 7.8 mm, cw 5.9 mm, bcw 5.8 mm) ( ZRC 2020.378 View Materials , ex MNHN-IU-2017-8792) , 1 male (cl 5.9 mm, pcl 4.6 mm, cw 4.2 mm, bcw 3.2 mm), 1 female (cl 7.5 mm, pcl 6.3 mm, cw 5.8 mm, bcw 4.5 mm) (MNHN-IU-2017-8792), stn CC3150 , Mozambique Channel , 1930’S 3646’E, 261–264 m, coll. MAINBAZA, 13 April 2009 :
Comparative material. Eurynome aspera ( Pennant, 1777) : 6 males (cl 18.1 mm, pcl 13.9 mm, cw 13.8 mm, bcw 10.3 mm; cl 15.7 mm, pcl 12.8 mm, cw 12.0 mm, bcw 9.1 mm; cl 14.7 mm 12.2 mm, cw 11.6 mm, cw 8.7 mm; cl 14.4 mm, pcl 11.5 mm, cw 10.7 mm, bcw 8.5 mm; cl 12.4 mm, pcl 11.6 mm, cw 11.5 mm, bcw 8.8 mm; cl 11.7 mm, pcl 9.3 mm, cw 8.9 mm, bcw 7.0 mm) ( ZRC 1988.665 View Materials - 670 View Materials ), Rovinj , Croatia, Mediterranean, on sedimentary bottom, 25 m, coll. Z. Števčić, 30 July 1986 .
Type locality: The Cape of Good Hope
Diagnosis. Pseudorostral spines short to long, outer margins; ventral surface concave ( Figs. 2A, B View FIGURE 2 , 3A, B View FIGURE 3 ). Supraocular eave with concave margin; postocular tooth forming cup in which eyes protected. Large hepatic tooth flattened, directed laterally outwards. Lateral border of carapace with 3 or 4 large boletiform granules. Gastric region with 3 larger granules, with rows of long hooked setae; lateral branchial region with 2 larger granule like-teeth; cardiac area with ring of flat granules around median larger one. Posterior border of carapace with 2 posteriorly directed teeth. Basal antennal article wide, with deep longitudinal groove ( Fig. 3B View FIGURE 3 ). Buccal frame quadrangular; third maxilliped with merus clearly separated from ischium, ischium covered by numerous rounded granules, with longitudinal groove ( Fig. 3D View FIGURE 3 ). Anterior part of male thoracic sternum depressed with prominent plates on margins of sternopleonal cavity surrounding telson ( Fig. 2B View FIGURE 2 ). Male cheliped long, covered by large granules ( Fig. 2A, D View FIGURE 2 ). Ambulatory legs relatively short with large granules only P2–4 meri, dorsal carina not continuous; dactylus proportionately shorter; carpi, propodi and dactyli covered by setae ( Figs. 2A View FIGURE 2 , 3C View FIGURE 3 ). G1 relatively shorter, gently curved, with large curved subdistal projection on interior border, hook-like ( Fig. 13D–F View FIGURE 13 ).
Remarks. Stimpson (1857: 220) described E. longimana from material collected from False Bay in the Cape of Good Hope, South Africa, but no figure was provided. Rathbun (1893: pl. 8 fig. 1) listed E. longimana in her catalogue but although she noted the USNM did not have specimens, she did provide a figure of the species. The figure is somewhat schematic and does not show the plates on the carapace or the tuberculation on the chelipeds clearly. Stimpson (1907: 27) figured the species again, noting that he had several specimens, the largest male being 11.9 by 8.6 mm, the largest female 9.9 mm in carapace length, all collected from 18 m in a rocky substrate among gorgonians. He described the fresh specimens as “dull red: feet whitish or variegated with pale red: eyes small black ( Stimpson 1907: 27).
Barnard (1950: 57) regarded E. aspera and E. longimana as synonymous, though he admitted he did not have comparative material from Europe. Capart (1951: 87) briefly discussed the identity of Barnard’s (1950) material and suggested that E. longimana may be distinct from E. aspera , a view which Monod (1956: 482) appear to share as he queried the presence of E. aspera in South Africa. Griffin (1974: 12), with material from Durban, discussed the matter of the two species again, arguing that the degree and pattern of the carapace tuberculation as well as setation of the G1 cannot discriminate between the two species, and continued regarding them as synonyms. He also noted that there is one extant type, a male syntype of E. longimana , c.l. 9.3 mm, from “ Cape Town, South Africa, North Pacific Exploring Expedition” in the collections of the Museum of Comparative Zoology, Cambridge (Mass.), U.S.A. The specimen preserved in spirit bears the registration number R50 (Griffin 1974: 12).
Comparisons of the material on hand of E. aspera from the Mediterranean and the European literature suggests that recognizing just one wide-ranging species is probably incorrect. While the structures of the carapace tubercles, pattern of tuberculation and shapes of the pseudorostrum are clearly unreliable characters and subject to variation, comparison of the present specimens from the Mozambique Channel with material from Croatia in the Mediterranean show five distinguishing features that suggest that we are dealing with a species complex here. The specimens from the Mediterranean have the outer surface of the ischium of the third maxilliped covered only with few large rounded granules, and are sometimes low in larger specimens ( Fig. 4B, F View FIGURE 4 ); the ambulatory merus is proportionately longer with at least the distal half of the dorsal margin prominently carinate ( Fig. 4A, D, E View FIGURE 4 ; Hartnoll 1961: fig. 1); the ambulatory dactylus is proportionately longer in E. aspera ( Fig. 4D, E View FIGURE 4 ; Hartnoll 1961: fig. 1); the triangular plate adjacent to the rimmed tip of the sternopleonal cavity is broadly triangular ( Fig. 4C View FIGURE 4 ; Hartnoll 1961: fig. 3a); and the main stem of the G1 structure (before the hook-like projection) is relatively more slender and longer ( Hartnoll 1961: fig. 1; Richer de Forges & Ng 2007: fig. 5A). This contrasts with the eastern African specimens, where the outer surface of the ischium of the third maxilliped is covered with numerous large rounded granules ( Fig. 3D View FIGURE 3 ); the ambulatory merus is proportionately shorter with the dorsal carina not distinct and not present as a continuous plate ( Figs. 2A View FIGURE 2 , 3C View FIGURE 3 ); the ambulatory dactylus is proportionately shorter ( Figs. 2A View FIGURE 2 , 3C View FIGURE 3 ); the triangular plate adjacent to the rimmed tip of the sternopleonal cavity is acutely triangular ( Fig. 2B View FIGURE 2 ); and the main stem of the G1 structure (before the hook-like projection) is relatively shorter ( Fig. 13C–E View FIGURE 13 ). Noteworthy is that the E. longimana figures of Rathbun (1893) and Stimpson (1907: pl. 4 fig. 2) both depict a species closely resembling the present material, notably in the relatively short ambulatory legs. These observations suggest that E. longimana should be recognised as a distinct species, at least until all the extant material can be compared and reassessed. Eurynome aspera was described from Port Erin in the United Kingdom (the types are no longer extant, Hartnoll 1961: 171), and its three remaining synonyms will need to be reviewed when the necessary revision is made: E. scutellata (from Mediterranean); E. boletifera (from Mediterranean), and E. aspera var. acuta (from Cape Verde).
The specimens of E. aspera from South Africa by Barnard (1950: 56, fig. 12a) from False Bay and Agulhas Bank differ slightly from the present material of E. longimana . In the present specimens, the pseudorostral spines are proportionately shorter and more flattened ( Figs. 2A View FIGURE 2 , 3A, B View FIGURE 3 ) (versus proportionately longer and less obviously dorsoventrally flattened; Fig. 13A View FIGURE 13 ; Barnard 1950: fig. 12a); the gastric, cardiac and branchial regions are more prominently raised and clearly demarcated ( Fig. 2A View FIGURE 2 ) (versus less well delimited; Fig. 13A View FIGURE 13 ; Barnard 1950: fig. 12a); and the branchial area is raised and carries strong spines (one branchial and two epibranchial) ( Fig. 2A View FIGURE 2 ) (versus with only small spines; Fig. 13A View FIGURE 13 ; Barnard 1950: fig. 12a). These differences do not appear significant and we treat them as variations of E. longimana for the time being. Their G1 structures are identical ( Fig. 13C–E View FIGURE 13 ). The west African material from Dakar reported by Monod (1956: 481) appear to be closer to E. longimana as defined here. The proportions of the ambulatory legs and G1 structure ( Monod 1956: 481, figs. 646, 647) certainly agree better with this species rather than E. aspera s. str.
Several species previously thought to be widespread across Europe and the eastern Atlantic that are present in the Indian Ocean via South Africa have been shown to be separate taxa, for example: Maja cornuta ( Fabricius, 1787) versus M. squinado ( Herbst, 1788) (Majidae) (cf. Ng & Richer de Forges 2015); Scyramathia carpenteri ( Thomson, 1873) versus S. hertwigi Doflein , in Chun, 1904 ( Epialtidae ) (cf. Lee et al. 2020); and Goneplax rhomboides ( Linnaeus, 1758) versus G. clevai Guinot & Castro, 2007 (Goneplacidae) (cf. Guinot & Castro 2007).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Eurynome longimana Stimpson, 1857
| Forges, Bertrand Richer De, Lee, Bee Yan & Ng, Peter K. L. 2021 |
Eurynome aspera
| Barnard, K. H. 1950: 56 |
Eurynome longimana
| Monod, Th. 1956: 482 |
| Capart, A. 1951: 87 |
| Lenz, H. & in Lenz, H. & Strunck, K. 1914: 275 |
| Stebbing, T. R. R. 1910: 289 |
| Stimpson, W. 1907: 27 |
| Rathbun, M. J. 1893: 102 |
| Stimpson, W. 1857: 220 |