
Eupithecia penumbrata (Pearsall)
|
publication ID |
https://doi.org/ 10.5281/zenodo.158660 |
|
publication LSID |
lsid:zoobank.org:pub:313752D4-D3AE-4F77-8D88-66867F938970 |
|
DOI |
https://doi.org/10.5281/zenodo.6271069 |
|
persistent identifier |
https://treatment.plazi.org/id/2220FE41-1116-B11F-EE65-FAAAF253FD38 |
|
treatment provided by |
Plazi |
|
scientific name |
Eupithecia penumbrata (Pearsall) |
| status |
|
Eupithecia penumbrata (Pearsall) View in CoL [New Synonomy] ( Fig. 16 View FIGURE 16 , HT)
Original description: Eucymatoge penumbrata Pearsall, R. F., 1912 . Can. ent. 44: 29.
Type series and location: Holotype female in AMNH; allotype male in NMNH.
Type locality: Palmerlee [Miller Canyon, Huachuca Mts.], Arizona; male allotype from Redington, Arizona, January 1, 1910.
McDunnough revision: page 676; Plate 31, fig. 28 female holotype; text fig. 18B male genitalia. Additional illustration: Barnes, W. & McDunnough, J. H., 1918. Contrib. nat. hist. Lepid. North America. p. 202, plate 22, fig. 8 male allotype. Color photo in Knudson & Bordelon (2003), pl. 6, fig. 3 ( Guadalupe Mts. Nat. Park section).
Diagnosis: Separated from other species by its elongate FW and relatively large size (FWL = 11.0–13.0 mm), DFW dark discal spot/dash with reddish center, heavilyscaled dark maculation, and antemedial transverse black bar (males and some females).
Discussion: McDunnough (1949) recognized the similarities between classicata and penumbrata , but chose to maintain them as separate species [as did Scoble (1999)] based upon the difference in the time of occurrence ( penumbrata early in the year and classicata in late summer) and the apparently smaller size of classicata relative to penumbrata in the few specimens he examined: classicata , HT, one additional male from Pinal Co., AZ; penumbrata , HT, allotype, one female in AMNH collection from Redington, AZ with the date February 1–10. McDunnough illustrated only the male genitalia.
Based upon material I examined, I see no differences in gross morphology or genitalia to support maintaining two separate species and hereby place penumbrata as a synonym of classicata , which has date priority. Apparently this view was expressed at some point in the past based upon the comments by parties unknown on a pin label of the penumbrata type ( Fig. 15 View FIGURE 15 ): “Is this a synonym of Eucymatoge classicata Pears. type in Grossbeck Coll.” with “yes” overwritten. There is no temporal separation since adults have been collected in all months of the year (no June record for Cochise Co., AZ). The HT of classicata ( Fig. 16 View FIGURE 16 ) is a male and the HT of penumbrata a female. Adults are sexually dimorphic with some degree of variation ( Fig. 17 View FIGURES 17 ); most females essentially lack the DFW antemedial diagonal black bar found in the males, and it is replaced by some thin striations; the bar was distinct in 2 of 25 females examined. The apparent smaller size of late summer adults is easily explained. Presumably the larvae, which are assumed to be flower or leaf bud feeders, matured during the normally very dry spring months when forage is limited, and were thus malnourished leading to small adults. Larvae maturing after the summer monsoon rains or the late winter wet period would have adequate food to achieve full growth. The nature of the male and female genitalia suggests that classicata is allied to E. graefii (Hulst) , but the two species differ substantially in wing shape and maculation. In both sexes there is a patch of ruddy scales located within the DFW black discal dash or spot, a character often seen in graefii .
Genitalic dissections: 3 males, 7 females. Male genitalia ( Figs. 18–19 View FIGURE 18 View FIGURE 19 ): Sclerotized plate on eighth sternite long, deeply dissected (18e, 19g), much more so than graefii , with two tines of unequal length. Symmetric valvae (18a, 19a).Terminal (segment IX) hair pencils well developed (18b, 19b). Vesica armature of aedoeagus (18c–d, 19d–f) consists of two robust unequal length pointed spines and a third contorted piece of chitin. Uncus tip forms a single point (similar to graefii ).
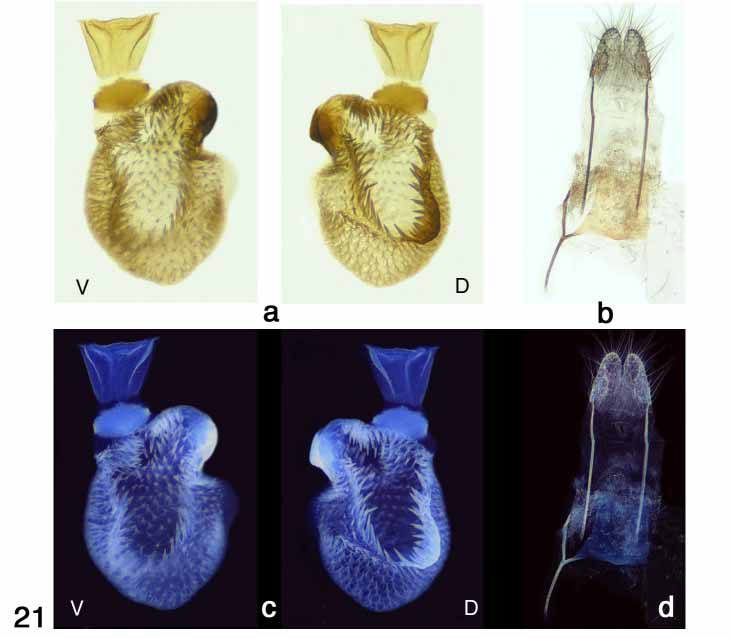
Female genitalia ( Figs. 20–21 View FIGURE 20 View FIGURE 21 ). General form similar to graefii , with ductus seminalis (20b–d, 21a) moderately stout at base, but tapering quickly and essentially transparent, rendering it difficult to visualize under the microscope and to photograph. Dorsal surface of corpus bursae displays a prominent ushaped array of robust spines attached to a chitinous base (20b:D, 21a, c:D).
Biology. Pupae found by Ron Wielgus on Arbutus arizonica , Upper Ash Canyon, late February and early March, 1986. In view of the date, locality, and altitude records for this species, there must be overlapping generations with various larval hosts. In the Huachuca Mts., classicata occurs in the same habitat as biedermanata ( Fig. 22 View FIGURE 22 ). It also occurs in open coniferous forest (Rustler Park, Chiricahua Mts.).
Geographic distribution: ARIZONA, Cochise & Pinal counties; OREGON, Baker Co. [one July record, Rindge (1963), possible misidentification of graefii ?]; TEXAS, Uvalde Co. (Dog Canyon); MEXICO, Durango.
Flight period: All year subject to climatic conditions. ARIZONA, Jan.–May, July– Dec.; TEXAS, Apr.–Sept.; MEXICO, Durango, August.
Material examined (17m, 25f; types by photos): ARIZONA, Cochise Co., Huachuca Mts.: Ash Canyon, early Sept.–midMay, 1981–2002, N. McFarland (9m, 8f), 17, 21.iv.2004, C. D. Ferris (4f), 1.x.2004, C. D. Ferris (1m); Copper Canyon, 1830m, 15.iv.86, J. Powell (1f); Lutz Canyon, 1800 m, 28.iv.89, J. Powell (2f); Miller Canyon, 14.iv.86, Powell & Wagner (1m, 1f), 1775 m, 1213.iv.88, J. Powell (3m, 3f); Townsite C.G., 7000’, 1.v.89, J. Powell (2m, 1f); Chiricahua Mts.: Rustler Park, 2590m, 2.viii.73, J. Powell & S. Szerlip (1f); 12.vii.72, J. Powell (1f). MEXICO, Durango, Las Rosias, 12 mi. E. La Ciudad, 2805 m, 1418.viii.72, J. Powell, D. Viers & C. D. MacNeill (1m, 1f); Tepalcates, 30 mi. W. Durango, 11.viii.86, Brown & Powell (1f); 10 mi. E. La Ciudad, Las Rusias Fire Lookout, 2805 m, 16.viii.72, J. Powell, D. Viers & C. D. MacNeill (1f).
Comment. Based on observations in Cochise Co., Arizona, Noel McFarland (pers. comm.) reports that under suboptimal conditions, Eupithecia pupae may remain dormant for several years until climatic conditions are suitable for emergence and survival of the subsequent larvae. Emergence seems to be triggered by increased humidity during the two seasonal rainy periods, midwinter and summer monsoon. Vegetation flowering occurs following these rainy periods and supports Eupithecia larvae that feed in flower heads. Following the winter rainy period, A. pungens (manzanita) blooms in late winter into early spring. Its flowers support the larvae of E. gilvipennata , hohokamae, and scabrogata. The mature larvae of these species are virtually identical, being cryptically patterned and colored to match the white to pink flowers. April flowering of A. arizonica (madrone) supports E. biedermanata larvae and apparently those of classicata .







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |