
Endecous (Endecous) zin, Carvalho & Junta & Castro-Souza & Ferreira, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5263.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3386FD59-2075-4F6A-8B21-B36C7F463EA9 |
|
DOI |
https://doi.org/10.5281/zenodo.7797747 |
|
persistent identifier |
https://treatment.plazi.org/id/B3578783-FFBE-E851-FF44-FDE6FCDF8C0F |
|
treatment provided by |
Plazi |
|
scientific name |
Endecous (Endecous) zin |
| status |
sp. nov. |
Endecous (Endecous) zin n. sp.
( Figures 32 – 36 View FIGURES 32–36 , 37 – 45 View FIGURE 37–45 , 46 – 47 View FIGURES 46–47 , 48 – 51 View FIGURES 48–51 , 52 – 57 View FIGURES 52–57 , 58 – 63 View FIGURES 58–63 , 64 – 66 View FIGURES 64–66 ; Table 2)
Material examined. Holotype, ♁, code ISLA 104845, Brazil, Bahia, municipality of Serra do Ramalho, Gruna das Três Cobras cave (43°45′9.87″W, 13°37′6.62″S), 15.ix.2021, R. L. Ferreira; condition: right tegmen, left legs and phallic complex were detached/dissected and stored alongside the holotype GoogleMaps . Paratypes, 1 ♀, ISLA 104846, municipality of Serra do Ramalho , Gruna das Três Cobras cave (43°45′9.87″W, 13°37′6.62″S), 26.viii.2022, R. L. Ferreira; GoogleMaps 5 ♁♁, ISLA 104847, 104848, 104849, municipality of Carinhanha, Gruna Grande cave (43°45′18.19″W, 13°36′8.9″S), 21.ix.2021, R. L. Ferreira; GoogleMaps ISLA 104850, municipality of Carinhanha, Gruna do Google cave (43°48′49.04″W, 13°37′41.93″S), 22.ix.2021, R. L. Ferreira; GoogleMaps ISLA 104851, municipality of Serra do Ramalho, Gruna da Serra Solta II cave (43°45′7.48″W, 13°30′38.16″S), 18.ix.2021, R. L. Ferreira GoogleMaps .
Etymology. The epithet “zin” refers to a figure of speech often used in Brazil to refer to a friend. It is actually a shortening of “Zezin”, which is a nickname for “José”, a very common name in Brazil.
Diagnosis. Combination of the following characteristics: pseudepiphallic dorsal branches (Ps.db) elongated, inclined towards the parameres, projected towards the dorsal portion of phallic complex, apex tapered and acute; pseudepiphallic ventral branches (A) elongated and dilated at the apex; pseudepiphallic inner bars (Ps.ib) concave; pseudepiphallic rami (R) developed, forming a membranous shield that partially covers the ectophallic apodeme laterally; ectophallic arc (Ect.arc) broad and trapezoidal in general shape; ectophallic lateral bars (Ect.lb) elongated, meandering, slightly inclined towards the interior of the phallic complex, apex resembling a hook; endophallic sclerite anterior portion (End.sc.a) with a crest on the opposite side of its central groove.
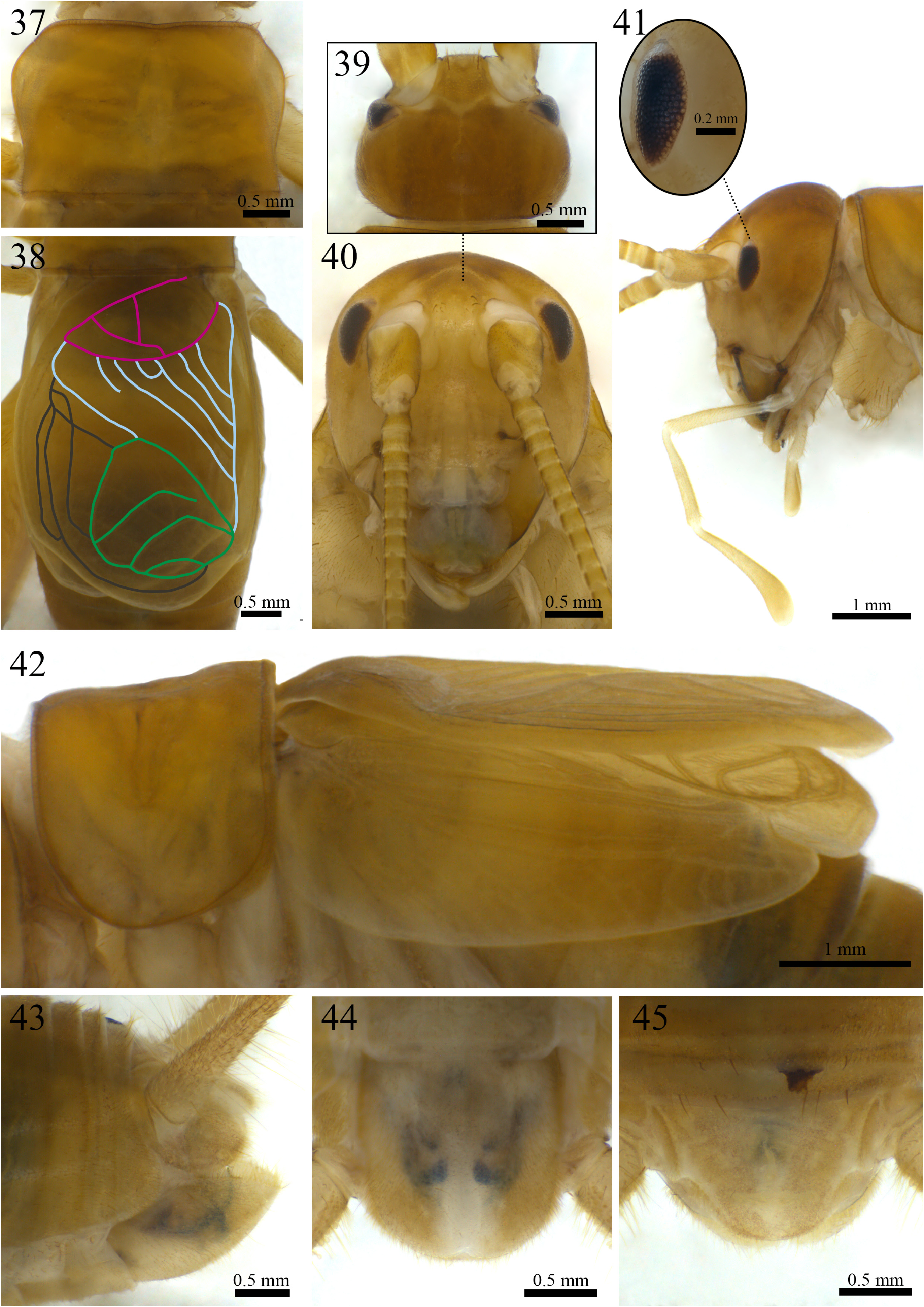
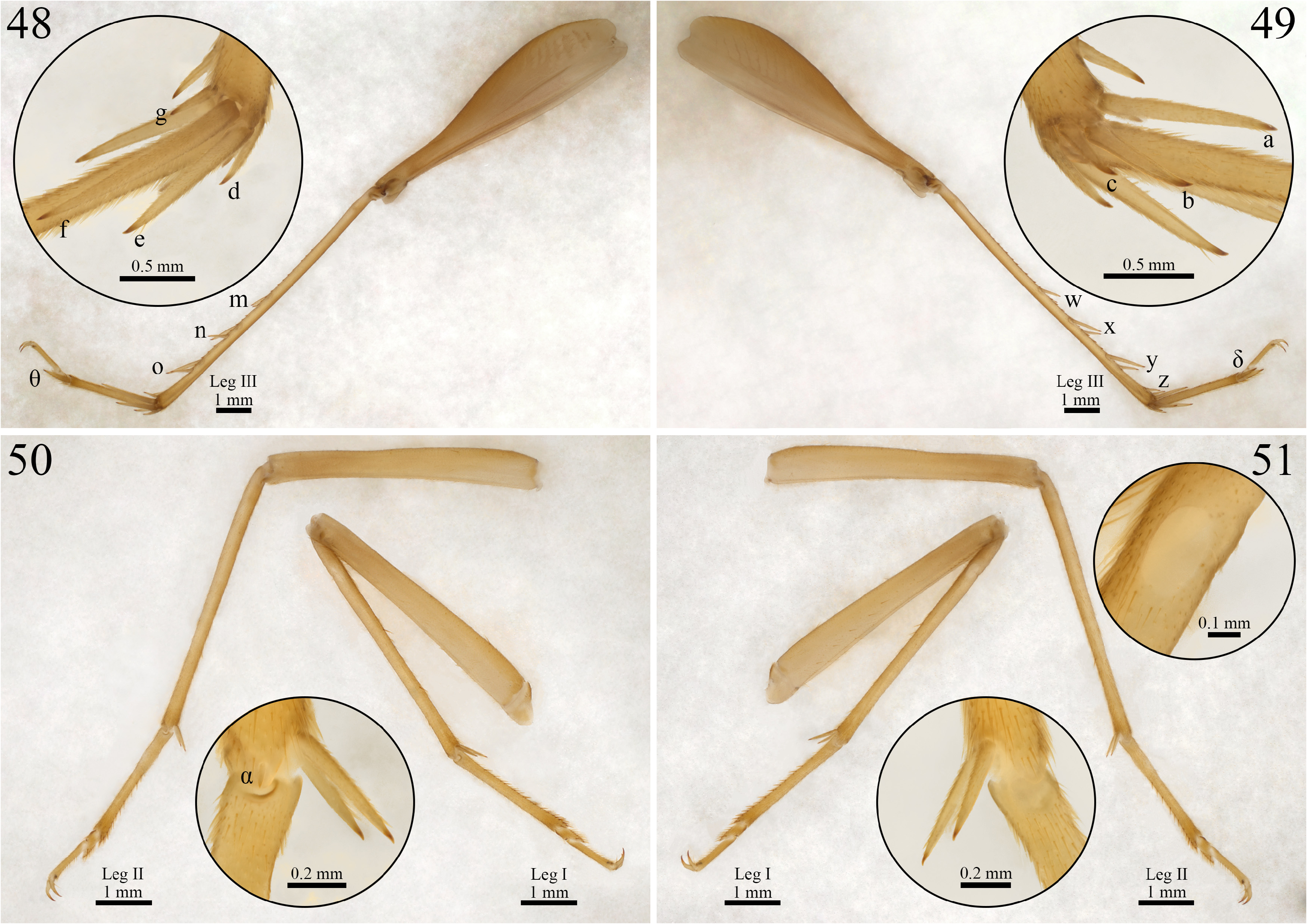
Morphology (holotype ISLA 104845, Figs. 37–45 View FIGURE 37–45 ). Body color: dorsal head yellowish brown; pronotum yellowish brown; abdomen whitish at its anterior portion and brownish at its posterior portion, and whitish ventrally (Figs. 37,39–45); entire legs yellowish brown, whitish at their proximal portion ( Figs. 48–51 View FIGURES 48–51 ); cerci yellowish brown throughout all their extension ( Fig. 43 View FIGURE 37–45 ). Head: slightly pubescent, elongated in frontal view; front yellowish brown; gena whitish brown; fastigium with long bristles, extending the vertex in an inclined plane pointing downwards; clypeus and labrum whitish with yellowish spots; mandibles yellowish brown and more sclerotized at the apex and margins; maxiles whitish and more sclerotized at the apex ( Fig. 40 View FIGURE 37–45 ); all maxillary palpomeres pubescent, the first two are whitish, short and same-sized, the other three palpomeres are longer and light yellowish brown, palpomere V (1.77 mm) is claviform and whitish at the apex (right maxillary palp missing) ( Fig. 41 View FIGURE 37–45 ); all labial palpomeres pubescent, increasing in size, palpomeres I and II are whitish, palpomere III is light yellowish brown and dilated from base to apex, which is whitish ( Fig. 41 View FIGURE 37–45 ); scape, pedicel and flagellomeres yellowish brown, distal portion whitish ( Fig. 41 View FIGURE 37–45 ); compound eyes reduced, ommatidia black; ocelli absent ( Fig. 41 View FIGURE 37–45 ). Thorax: pronotum lateroanterior and lateroposterior regions with long bristles; dorsal disk broader than long (1.94 and 2.86 mm in length and width, respectively), lateral lobes rounded and leaning towards the anterior portion of the body, anterior and posterior margins arched and sub-straight ( Fig. 37 View FIGURE 37–45 ). Legs: femur, tibia and tarsus pubescent ( Figs. 48–51 View FIGURES 48–51 ). Leg I ( Figs. 50 and 51 View FIGURES 48–51 ): tibia slightly longer than the femur, with an oval tympanum on its inner side and two same-sized ventral apical spurs, first tarsomere ventrally serrated and longer than the second and third tarsomeres together. Leg II ( Figs. 50 and 51 View FIGURES 48–51 ): tibia slightly longer than the femur, with two same-sized ventral apical spurs, first tarsomere ventrally serrated and longer than the second and third tarsomeres together. Leg III ( Figs. 48 and 49 View FIGURES 48–51 ): femur developed; tibia slightly longer than the femur (8.75 and 8.17 mm, respectively); tibia armed with four subapical spurs on the outer side, the distal being the shortest, and three on the inner side, the proximal being the shortest, three apical spurs on the outer side ( Fig. 49 View FIGURES 48–51 ; a, b, c), spur “a” being the longest and “c” the shortest, and four on the inner side ( Fig. 48 View FIGURES 48–51 ; d, e, f, g), spurs “e” and “f” longer than the “d” and “g”; first tarsomere longer than the second and third tarsomeres together, with two apical spurs, the inner one being the longest (right leg missing). Right tegmen: slightly sclerotized; covering the first four urotergites (5.12 and 3.39 mm in length and width, respectively); harp with four well-marked crossveins and five cells, a fifth transversal crossvein is visible, but short, and does not reach the opposite margin of the harp, one of the main crossveins derives from two converging crossveins; mirror subtriangular, with three well-marked crossveins and three cells, the most anterior vein does not reach the opposite margin of the mirror; basal field with a single bifurcated secondary vein connecting Cu2 to 2A, 1A absent; lateral field with two connected longitudinal veins and several irregular and weakly marked secondary veins ( Fig. 38 View FIGURE 37–45 ); stridulatory file with 54 teeth. Abdomen: cerci pubescent, with globose setae at the base, mainly on the inner side, and long bristles throughout all their extension (right cerci broken); sub-genital plate whitish brown, distal margin rounded, with a small dent in the center ( Fig. 44 View FIGURE 37–45 ); supra-anal plate yellowish brown, shorter than the sub-genital plate, subtriangular, distal margin rounded, lateral projections short and curved outwards ( Fig. 45 View FIGURE 37–45 ); paraprocts as long as the supra-anal plate and barely visible in dorsal view ( Fig. 45 View FIGURE 37–45 ).
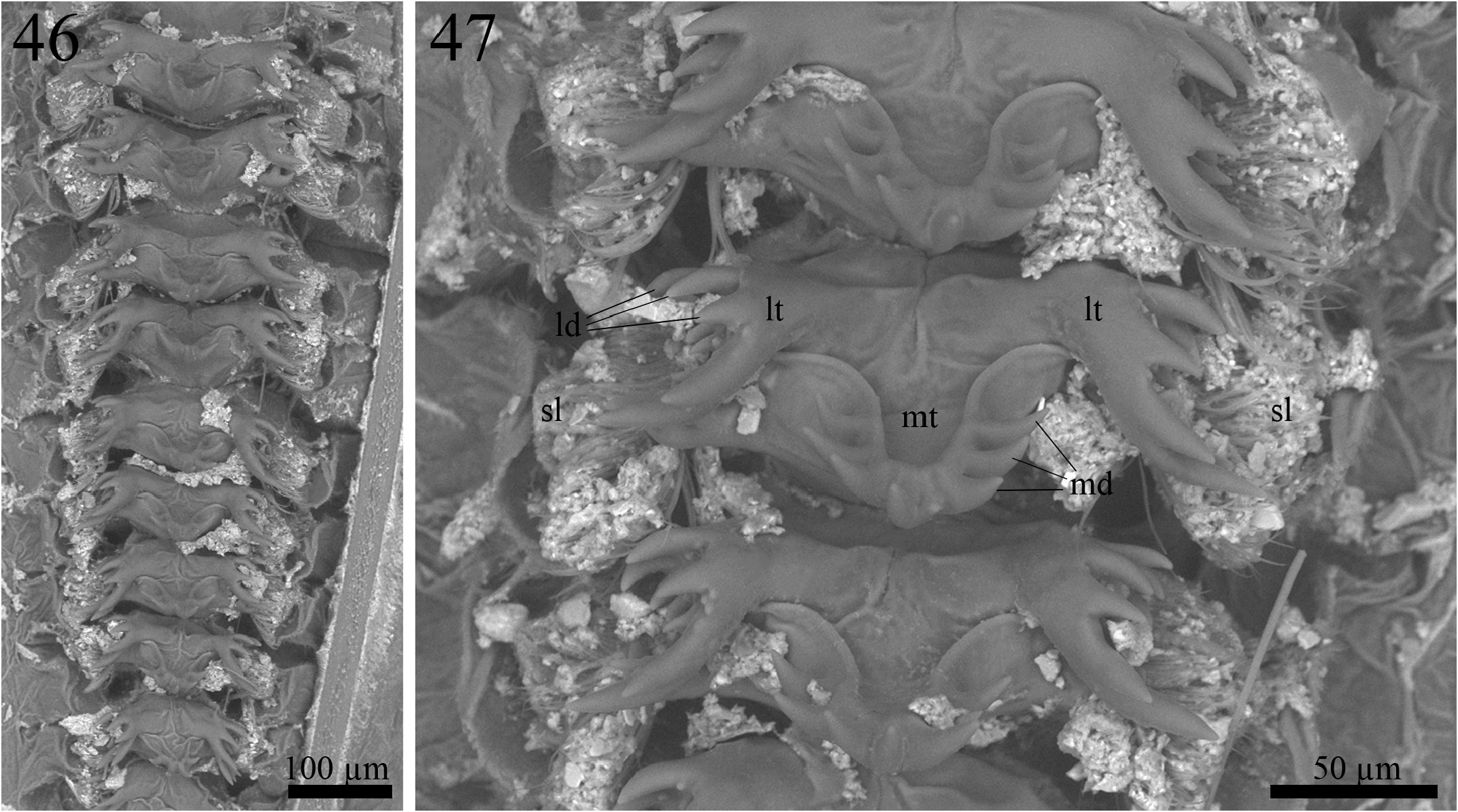
Proventriculus (paratype ISLA 104848, Figs. 46–47 View FIGURES 46–47 ). Proventriculus internally organized in six rows of 13 overlaid sclerotized appendices (sa); sclerotized lobes (sl) bearing a cluster of bristles are visible on each side of the sclerotized appendices; sclerotized appendices formed by a median tooth (mt), with at least five, but up to seven median denticles (md), and two lateral teeth (lt), with several lateral denticles (ld); a single median tooth showed eight median denticles; median denticles elongated, tapered and acute at the tip; lateral denticles also elongated, tapered and acute at the tip; the most posterior sclerotized appendix does not have a median tooth.
Male phallic sclerites (holotype ISLA 104845, Figs. 32–36 View FIGURES 32–36 ). Phallic complex broadened at the proximal and central portions, with an almost trapezoidal contour in dorsal and ventral views ( Figs. 32 and 33 View FIGURES 32–36 ). Pseudepiphallus: arms short and slightly inclined inwards in dorsal view ( Fig. 32 View FIGURES 32–36 , Ps.arm); dorsal branches long, dorsally projected, tapered and acute at the apex, which is slightly inclined towards the interior of the phallic complex in frontal view ( Fig. 35 View FIGURES 32–36 , Ps.db); “A” sclerite well-developed, elongated and dilated at the apex ( Figs. 34 and 36 View FIGURES 32–36 , A); paramere 1 and 2 fused in a single circular and concave structure that does not extend itself past the dorsal branches in dorsal view, half-moon-shaped in frontal view, distal region highly sclerotized, dorsal portion sclerotized, forming a circular edge that reaches the tip of the posterior portion of the ectophallic median projections in dorsal view ( Fig. 32 View FIGURES 32–36 , Ps.p); inner bars concave and slightly curved downwards in dorsal view ( Fig. 32 View FIGURES 32–36 , Ps.ib); rami developed, forming a membranous shield that partially covers the ectophallic apodeme in lateral view ( Fig. 36 View FIGURES 32–36 , R). Ectophallic invagination: arc well-developed, wide and trapeze-like in ventral view ( Fig. 33 View FIGURES 32–36 , Ect.arc); apodeme developed and slightly inclined towards the interior of the sclerite in dorsal view ( Fig. 32 View FIGURES 32–36 , Ect.ap); lateral bars developed, sinuous, longer than the ectophallic median projections, slightly inclined inwards, with a hook-shaped apex in ventral view ( Fig. 33 View FIGURES 32–36 , Ect.lb); median projections long, slender, slightly curved towards the exterior of the sclerite and reaching the parameres in dorsal view ( Fig. 32 View FIGURES 32–36 , Ect.mp). Endophallus: anterior portion well-developed and sclerotized, with a central groove and a crest opposite to the groove in lateral view ( Fig. 36 View FIGURES 32–36 , End.sc.a.); duct short, membranous, barely exceeding the length of the ectophallic arc in ventral view ( Fig. 33 View FIGURES 32–36 , End.sc.d); posterior portion membranous, trapezoidal at the apex, exceeding the length of the ectophallic median projections considerably and almost reaching the pseudepiphallic dorsal branches at their apexes in dorsal view ( Fig. 32 View FIGURES 32–36 , End.sc.p).
Variations in phallic sclerites (paratypes, n = 5, ISLA 104847, 104848, 104849, 104850, 104851). Phallic complexes vary slightly in size and degree of sclerotization; pseudepiphallic arms (Ps.arm), although always inclined to the interior of the sclerite, may vary slightly in the degree of inclination; right and left pseudepiphallic dorsal branches (Ps.db) with misaligned apexes as they extend dorsally and point to the center of the phallic complex; pseudepiphallic inner bars (Ps.ib) may be more or less concave, inclined or not towards the anterior and central part of the sclerite; ectophallic apodeme (Ect.ap) may or may not be inclined to the interior of the phallic complex; the distance between the endophallic sclerite posterior portion (End.sc.p) and the apex of the pseudepiphallic dorsal branches differs; in addition to that, the shape of the endophallic sclerite posterior portion also varies slightly, but still maintains the trapezoidal outline.
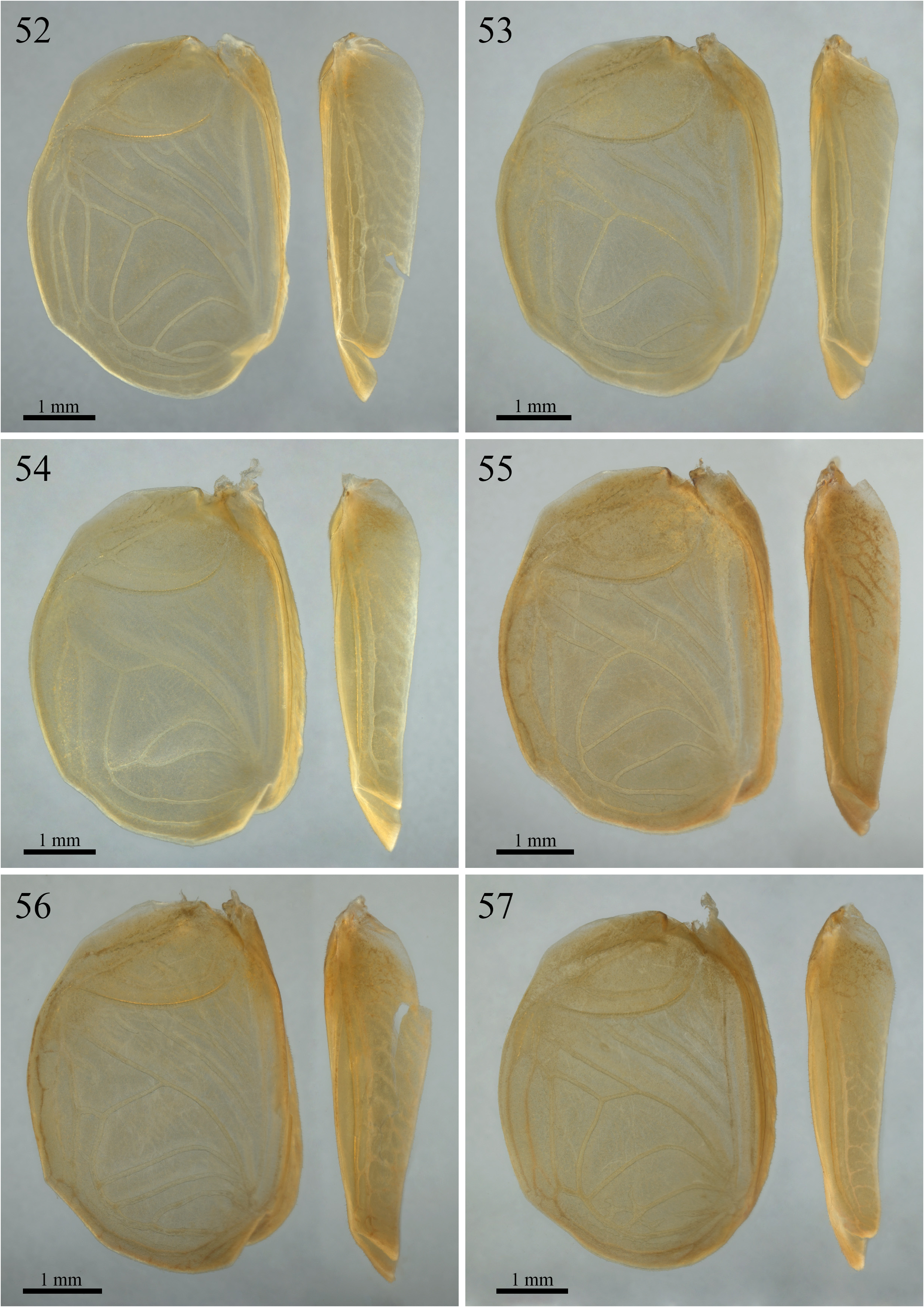
Variations in male right tegmen ( Figs. 52–57 View FIGURES 52–57 ). Stridulatory file with 56.17 ± 1.95 teeth (n° = 6, holotype and paratypes). Harp outline conserved; three to five diagonal crossveins; four to six cells; a short and incomplete diagonal crossvein may be present ( Figs. 55–57 View FIGURES 52–57 ). Mirror outline varies from subtriangular to almost arch-like; two or three arched crossveins; three or four cells; incomplete crossveins may be present ( Fig. 52 View FIGURES 52–57 ); irregular and poorly marked veins may also be present ( Fig. 57 View FIGURES 52–57 ); two main crossveins may be derived from a single bifurcated crossvein ( Fig. 54 View FIGURES 52–57 ). Basal field outline conserved; 1A may be poorly marked, incomplete ( Fig. 52 View FIGURES 52–57 ) or absent ( Fig. 53 View FIGURES 52–57 ). Lateral field with two parallel longitudinal veins that may or may not be connected.
Female (paratype ISLA 104846, Figs. 58–63 View FIGURES 58–63 ). Same coloration of the male, except for the dorsal side of the abdomen, which is, in this case, yellowish brown; apterous; bigger than the male (17.03 and 13.47 mm ± 1.02 mm in length, respectively); supra-anal plate yellowish brown, pubescent, elongated, conical in shape, with two small lateral projections, distal margin rounded and covered by long bristles ( Fig. 59 View FIGURES 58–63 ); sub-genital plate brown, pubescent, short, but broad, distal margin rounded, with a large V-shaped dent in the center ( Fig. 58 View FIGURES 58–63 ); ovipositor shorter than the cerci (7.5 mm), with a dorsoventral constriction near the apex, shaped like a curved sword ( Figs. 60 and 62 View FIGURES 58–63 ). Copulatory papilla sclerotized and short, anterior portion membranous, with an almost rectangular contour in dorsal and ventral views; distal portion widened, chalice-like, V-shaped ventral end longer than the also V-shaped dorsal end in lateral and dorsal views ( Fig. 63 View FIGURES 58–63 ).
Ecological remarks. Specimens of Endecous zin n. sp. were found in four caves located in the municipalities of Carinhanha and Serra do Ramalho, southwestern Bahia State, Brazil. The Gruna das Três Cobras cave was defined as the type locality. This cave has 5,620 meters of horizontal projection ( Silva et al. 2019), presenting internal conduits and chambers of labyrinthic aspect ( Fig. 64 View FIGURES 64–66 ). The cave has many entrances, most of them located at the base of the limestone outcrop. The cave atmosphere is constantly influenced by the airflow coming from those entrances and, as a consequence, several conduits dry out during the dry season ( Fig. 65 View FIGURES 64–66 ). However, due to the cave extension, there are moistened inner chambers located far from the entrances that remain humid even during the dry season. Specimens of Endecous zin n. sp. were observed in several areas along the cave, but never close to the entrances. They were usually walking on the cave floor or walls ( Fig. 66 View FIGURES 64–66 ). It is important to mention that this cave is partially flooded during the rainy season. Hence, the habitat of this species seems to be seasonally altered, thus, the crickets likely need to migrate to distinct areas in each season. The main organic resources occurring in the cave are plant debris from the epigean vegetation, and bat guano, the latter being more common in deeper regions of the cave. Such guano piles certainly represent an important food resource for this species, along with other decaying material, considering its generalist detritivore nature. This cave receives no human visitors, with the exception of few speleologists who sporadically explore it. Consequently, the cave is relatively well preserved. Nonetheless, the external area is highly degraded, with drone images revealing the extension of the logging activity in the cave surroundings ( Fig. 64 View FIGURES 64–66 ). Most areas were converted to pastures or had the vegetation removed and burned, maybe for future expansion of the grazing areas. As previously mentioned, such practices influence the organic input to the cave environment directly and, at the same time, contribute to the silting of many conduits and chambers inside the cave. Because of that, several microhabitats that might be important to this new species (among others) may be being severely altered by the external deforestation. In that sense, urgent actions are needed in order to prevent further negative impacts in the cave surroundings.
Endecous zin n. sp. was also observed in other three caves located not far from the Gruna das Três Cobras cave (Gruna do Google, Gruna Grande and Gruna da Serra Solta II caves). Such caves are considerably long (more than 1km of horizontal projection) and have intermittent drainages along their main conduit. The Gruna do Google and Gruna Grande caves were only visited by speleologists since its discovery, due to the difficulty in accessing their inner portions. Hence, its interior is quite preserved. The Gruna da Serra Solta II cave, on the other hand, is partially impacted, especially at the entrance area, where locals pump water from a cistern using a diesel engine. However, since the cave is quite long, most of its extension is devoid of visible impacts caused by humans. It is worth noting that there are many other caves in that area that were not sampled, so it is quite plausible to assume that E. zin n. sp. is more widely distributed in the region.
Similar to the Gruna das Três Cobras cave surroundings, the external landscape of Gruna do Google, Gruna Grande and Gruna da Serra Solta II caves is also altered, due to replacement of the original vegetation by pastures and crops. In that sense, it is important to highlight that the region is under risk, especially considering agriculture expansion and the growing threats of limestone extraction, which may impact the entire Serra do Ramalho region, affecting several caves, including those previously described.



| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Grylloidea |
|
Family |
|
|
SubFamily |
Luzarinae |
|
Genus |
|
|
SubGenus |
Endecous |