
Emphytopsis vernalis Shinohara
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3884.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:2F61B0CD-9FBA-42CB-958D-28008232F2BF |
|
DOI |
https://doi.org/10.5281/zenodo.6144110 |
|
persistent identifier |
https://treatment.plazi.org/id/039187A6-8A48-FFBF-5198-F95456B74666 |
|
treatment provided by |
Plazi |
|
scientific name |
Emphytopsis vernalis Shinohara |
| status |
sp. nov. |
Emphytopsis vernalis Shinohara , n. sp.
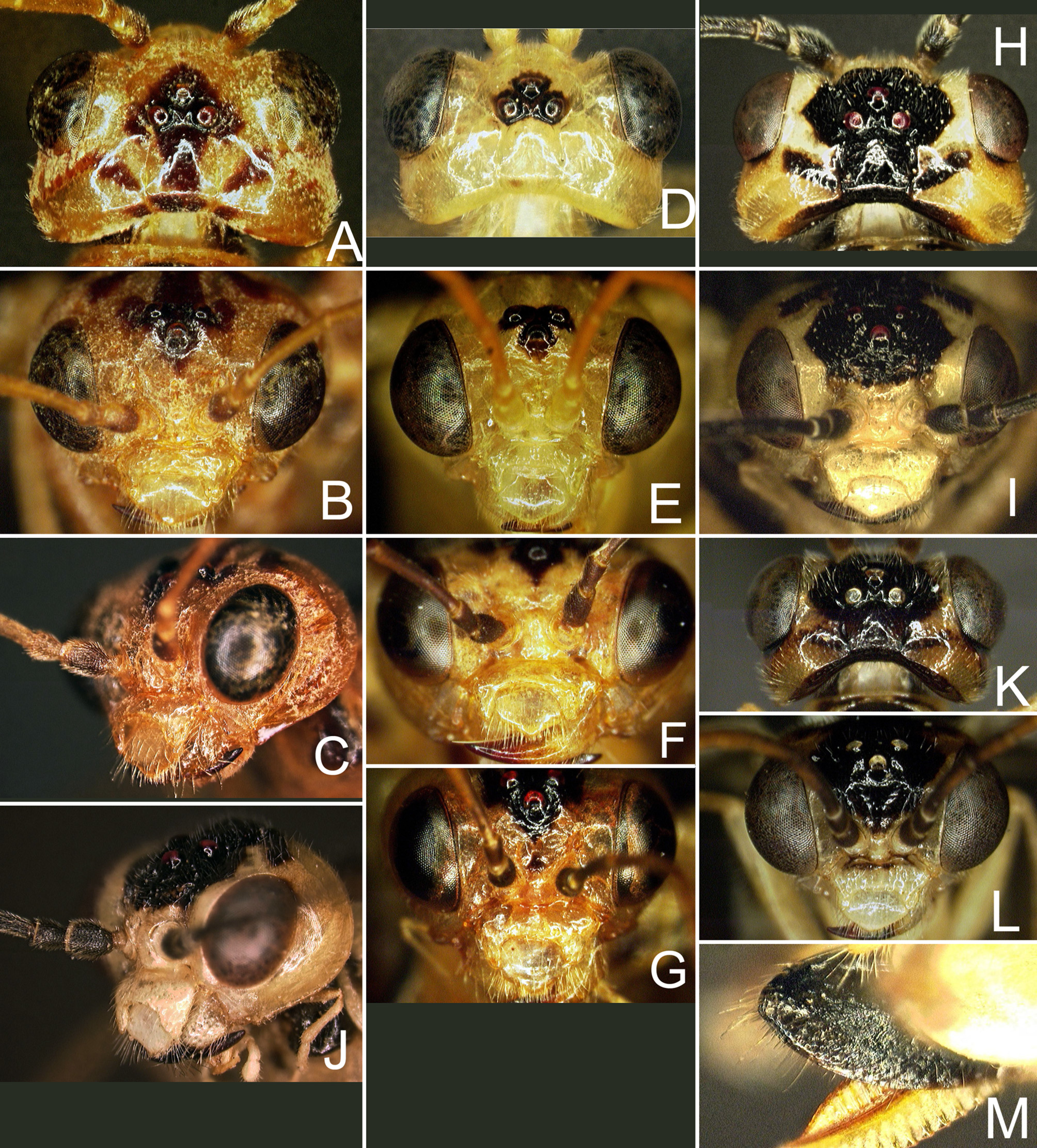
( Figs. 1 View FIGURE 1 F–K, 2G–L, 3H–M, 7, 8, 9E–H, L–N, 10)
Female (holotype). Length 9.5 mm. Brownish white (pale olive green in life) with black marks ( Figs. 1 View FIGURE 1 K, 2G–I). Head with large spot on upper surface, covering frons and large part of vertex, and most of occiput black ( Fig. 3 View FIGURE 3 H–J). Entire antenna and mandible (except for base) black. Pronotum with elongate mark along anterodorsal margin (medially interrupted) and large posteroventral spot black; propleuron and median part of prosternum black; mesoscutum black with whitish marking along borders of median and lateral lobes; mesopostnotum black; mesopleuron black, except for dorsal and median part of mesepisternum; metascutum and metapostnotum black; metapleuron black, except for large part of metepisternum. Legs brownish white with following black: Apical parts of fore and mid femora dorsally, apical parts of fore and mid tibiae, apical parts of fore and mid tarsomeres 1 and all other fore and mid tarsomeres (but not claws), apical 1/3 of hind femur, hind tibia (except for narrow base and basal dorsal surface), hind tarsus (except for very narrow base of tarsomere 1 and claws). Wings hyaline, very feebly stained with blackish brown; veins and stigma blackish brown; veins C and 1A and stigma basally narrowly pale. Abdomen black above, except for pale median longitudinal line; apical ovipositor sheath black.
Dorsal and lateral parts of head smooth, with very sparse indistinct punctures; frontal and supraclypeal areas slightly rugose; paraantennal field coriaceous, with dense minute punctures; clypeus rather densely punctured, interspaces smooth. Dorsum of thorax with rather dense, small punctures, interspaces smooth; sunken areas of mesoscutum coriaceous, without punctures; mesoscutellum and its appendage densely punctured with feebly shiny interspaces; metascutellum densely punctured and transversely microsculptured; metapostnotum polished; mesepisternum generally smooth, shallowly rugose dorsally, with rather dense small punctures; mesepimeron impunctate, dorsal part somewhat rugose; metapleuron mostly shallowly rugose. Abdominal segments coriaceous, feebly shining.
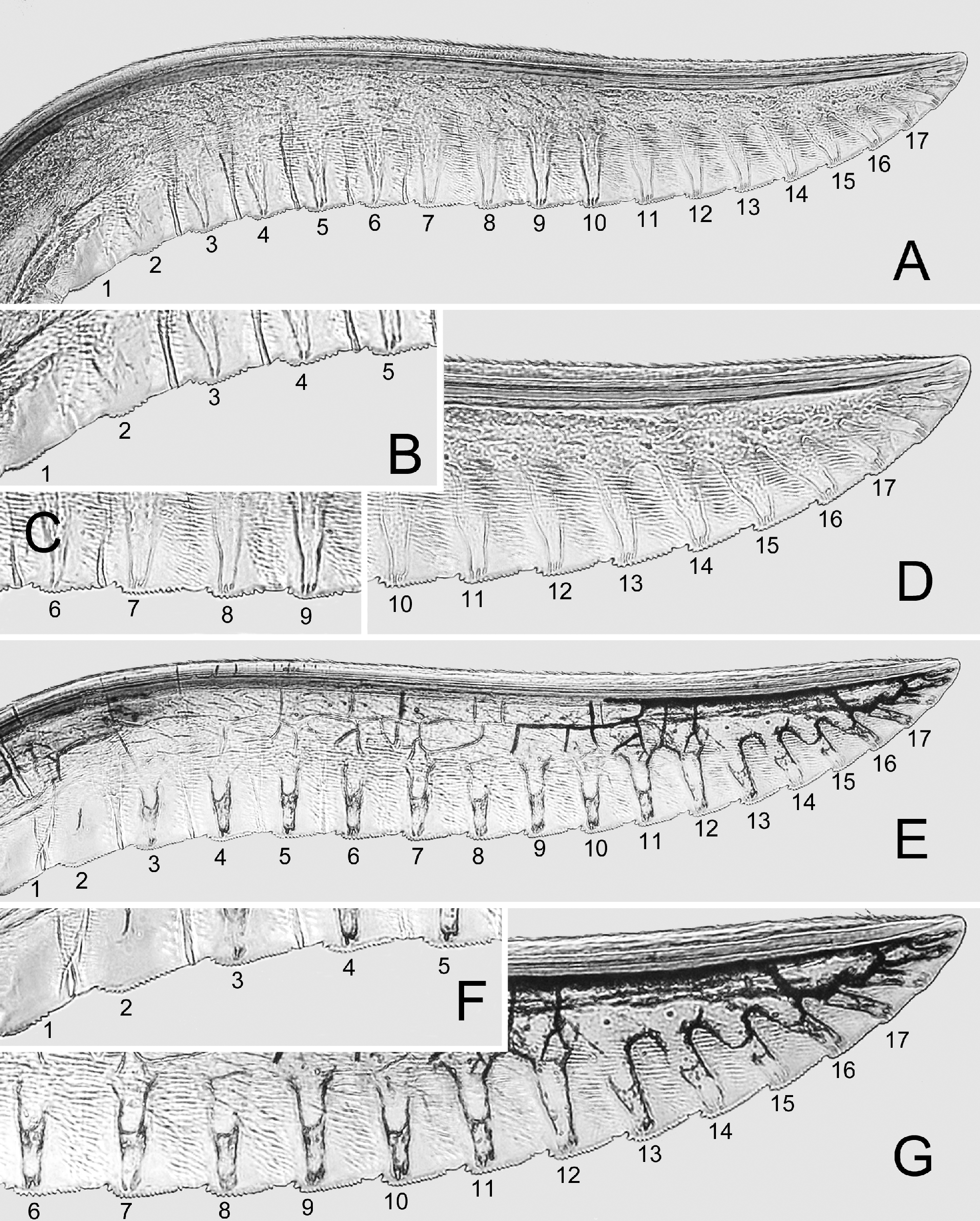
Anterior margin of labrum triangularly protruding ( Fig. 3 View FIGURE 3 I); anterior margin of clypeus incised to 0.43× clypeal length, lateral lobes rather stout, rounded at apex; malar space about 0.5× diameter of median ocellus; postocellar area weakly elevated, about as long as wide, lateral furrows weakly curved, subparallel, each with posterior half distinctly deepened to form longitudinal pit ( Fig. 3 View FIGURE 3 H); head behind eyes in dorsal view about as long as eye, lateral sides roundly convex, about as wide as across eyes. Antenna hardly tapering toward apex, relative lengths of antennomeres 1–9 about 7: 4: 14: 11: 9: 7: 6: 6: 6 (Fig. 2G–I). Mesoscutellum slightly convex in lateral view, with anterior slope shorter than posterior slope. Hind tarsomere 1 about 0.85× length of tarsomeres 2–5 together (Fig. 2G–I). Ovipositor sheath about 1.4× length of hind tarsomere 1, apical sheath about as long as basal sheath; apical sheath with both dorsal and ventral margins rounded and apex roundly produced ( Fig. 3 View FIGURE 3 M). Lancet ( Figs. 7 View FIGURE 7 , 8 View FIGURE 8 ) with about 19 serrulae, membrane between serrulae weakly convex; each serrula basally convex and gradually lowered apically, with 8 to 20 minute denticles.
Male (paratype from Nakagawa). Length 8.5 mm. Brownish white (pale olive green in life) with black marks (Fig. 2J–L). Head with large spot on upper surface, covering frons and large part of vertex, and most of occiput black ( Fig. 3 View FIGURE 3 K, L). Antenna (except for pale brownish ventral side of flagellum) and mandible (except for base) black. Pronotum with elongate mark along anterodorsal margin and large posteroventral spot black; propleuron except for pale inner ventral margin and median part of prosternum black; mesoscutum black with whitish marking along borders of median and lateral lobes; mesoscutellum brownish white, with anteromedian part black; mesopostnotum black; mesopleuron black, except for dorsal and median parts of mesepisternum; metascutum, metapostnotum and metapleuron black; metascutellum anteriorly black and posteriorly brownish white. Legs brownish white with following black: Narrow apical parts of fore and mid femora dorsally, apical part of fore tibia, apical and ventral parts of mid tibia, apical part of fore tarsomere 1 and all other fore and mid tarsomeres (but not claws), wide dorsal part and apical 1/2 of hind femur, hind tibia, hind tarsus (except for very narrow base of tarsomere 1 and claws). Wings hyaline, very feebly stained with blackish brown; veins and stigma blackish brown; veins C and 1A and stigma basally narrowly pale. Abdominal terga black, except for brownish white median longitudinal line and lateral margins (on ventral side); sterna entirely brownish white; genitalia mostly blackish brown to black.
Structure and punctuation generally similar to female. Anterior margin of labrum nearly rounded ( Fig. 3 View FIGURE 3 L); anterior margin of clypeus incised to 0.5× clypeal length; malar space about 0.2× diameter of median ocellus; postocellar area length about 0.9× width ( Fig. 3 View FIGURE 3 K); head behind eyes in dorsal view much shorter than eye, lateral sides roundly convex, much narrower than across eyes; relative lengths of antennomeres 1–9 about 6: 4: 13: 9: 8: 6: 6: 5: 6; hind tarsomere 1 about 0.72× length of tarsomeres 2–5 together (Fig. 2J–L). Subgenital plate very broadly rounded with nearly truncate apex. Genitalia as in Fig. 9 View FIGURE 9 E–H, L–N; harpe broad, with inner basal part strongly produced; valviceps in dorsal view ( Fig. 9 View FIGURE 9 E, F) with outer apical margin almost angled, and in lateral view ( Fig. 9 View FIGURE 9 G, H, L–N) with ventral margin rather strongly convex and thus anteroventral and posterodorsal margins subparallel.
Larva. Five molts before becoming prepupa in reared specimens of unknown sex. Last feeding instar (probably fifth) ( Fig. 1 View FIGURE 1 F): About 17–20 mm long; head pale brown with round black spot including eye; trunk entirely greenish white; whole insect covered with rather thick wax and consequently appearing whitish. Prepupa ( Fig. 1 View FIGURE 1 G, H) similar to last feeding instar but shorter, about 13 mm long, and more vivid in color and with no wax covering; a few basal thoracic segments inconspicuously darkened dorsally.
Variation. Female: The length ranges from 7.5 to 9.5 mm. The coloration is quite stable in the Tochigi specimens examined. The only female from Kyushu is darker; the apices of clypeal lateral lobes are blackish, the mesoscutum is entirely black, and the mesopleuron is entirely black, except for the large pale posterodorsal spot on the mesepisternum. The length/width ratio of the postocellar area ranges from 0.85 to 1 (average 0.92) and the ratio of the depth of ventral incision of the clypeus to the entire length of the clypeus ranges from 0.36 to 0.50 (average 0.42). The length of the malar space is about 0.4–0.6× diameter of the median ocellus. The length of the tarsomere 1 is about 0.82–0.91× lengths of the tarsomeres 2–5 together. Male: The length ranges from 6.5 to 9 mm. The basic color pattern is stable. Very dark specimens have the mesoscutum, mesoscutellum and metascutellum mostly black and the ventral sides of the abdominal terga are blackish. The length/width ratio of the postocellar area ranges from 0.87 to 1 (average 0.92) and the ratio of the depth of ventral incision of the clypeus to the entire length of the clypeus ranges from 0.38 to 0.52 (average 0.46).
Specimens examined. Type material: Holotype: ♀, Bambi Farm, 230 m alt., 3646’58 N 14010 View Materials ’29E, Wami, Nakagawa, Tochigi Pref., Honshu, 29. IV. 2010, Malaise trap, S. Ibuki. Paratypes: HONSHU—Tochigi Pref.: 1♂, Mino, 170 m alt., Kanuma, one of 13 larvae coll. 15. VI. 2008, mat. 16–23. VI., em. 4. IV. 2009, host: Stewartia pseudocamellia, T. Saito ; 1♂, same data but em. 5. IV. 2009; 1♂, same data but em. 6. IV. 2009; 1♂, same data but em. 7. IV. 2009; 1♂, same data but em. 10. IV. 2009; 1♂, same data but em. 12. IV. 2009; 3♂, same data but em. 13. IV. 2009; 4♀ 4♂, same locality, on Stewartia pseudocamellia , 12. IV. 2009, T. Saito; 10♀ 9♂, same data but 18. IV. 2009; 2♀ 2♂, same data but 26. IV. 2009; 4♀ 7♂, same data but 21. IV. 2012; 2♀ 6♂, Itaga, 230 m alt., Kanuma, 26. IV. 2011, T. Saito; 1♂, Bambi Farm, 230 m alt., 3646’58 N 14010 View Materials ’29E, Wami, Nakagawa, larva coll. 10. VI. 2011, mat. 11. VI., em. 16. IV. 2012, host: Stewartia pseudocamellia, S. Ibuki ; 2♂, same locality, 12. IV. 2012, Malaise trap, S. Ibuki; 2♀ 6♂, same data but 15. IV. 2012; 3♀ 4♂, same data but 16. IV. 2012; 14♀ 28♂, same data but 18. IV. 2012; 3♀ 1♂, same data but 1. V. 2012; 2♀, same data but 5. V. 2012; 1♀, same data but 14. V. 2012; 1♀ 5♂, same data but 10. IV. 2013; 5♀ 7♂, same data but 12. IV. 2014; 4♀ 4♂, same data but 13. IV. 2014; 2♀ 2♂, same data but 17. IV. 2014. Hyogo Pref.: 1♂, Kiritaki, Mt. Oginosen, Shin-onsen, 13. V. 2013, T. Naito. KYUSHU—Nagasaki Pref.: Mt. Gokaharadake, 1050m alt., Tarasan Mts., 16. V. 1997, A. Shinohara.
Distribution. Japan (Honshu, Kyushu).
Etymology. The species epithet, vernalis , refers to the relatively early, spring appearance of the adults of the new species.
Host plant. Theaceae : Stewartia pseudocamellia Maxim.
Field observations and rearing records. Saito observed feeding larvae on Stewartia pseudocamellia in Mino (170 m alt.), Kanuma City, Tochigi Prefecture, in 2007 but no attempt was made to rear them. On June 15, 2008, Saito collected 13 larvae in the same site. All of them stopped feeding, executed extra molt, thus becoming prepupae, and entered the soil during the period from June 16 to 23. In the soil, the prepupa made an earthen cell and stayed there. A total of nine male adults emerged from April 4 to 13, 2009 (see the list of specimens above).
In Mino, adults were found visiting the host plant on April 12, 2009, and many adults were observed in the same site on April 18, when the adults were frequently found on the leaves of Euonymus japonicus Thunb. ( Fig. 1 View FIGURE 1 K), possibly licking secretions of the larvae of a zygaenid moth, Pryeria sinica Moore, 1877 . On April 26, only a few adults were found, probably because the adult emergence season was close to the end. Eggs and some young larvae were also found and collected. On May 9, no adults were found but feeding larvae were common. A few of the larvae collected on April 26 matured on May 23. On June 11, several additional larvae were collected in the same site and they became prepupae and entered the soil on June 13–27. They all died during the hibernation.
On May 5, 2010, some feeding larvae were found but no adults. On April 26, 2011, many adults occurred on very new spring leaves of the host plant; Saito collected two females and six males.
On April 21, 2012, many adults, but no eggs, were found at the same site. A female adult was collected and kept alive in a cage with small twigs of the host plant. On April 29, the female died and many eggs were found on the leaves in the cage. Twenty-one eggs hatched on May 1. On May 4, no adults were found at the site in Mino but some young larvae were found.
Saito reared the larvae hatched on May 1, 2012, in six containers (A–F) as follows.
A (one larva): Two exuviae were found in the container on May 7, one exuvia each was collected on May 10, 14
and 21, respectively. On May 21, the larva (prepupa) went into the soil.
B (one larva): One exuvia each was collected on May 7, 8, 11, 16 and 23, respectively. On May 23, the larva
entered the soil.
C (one larva): One exuvia each was collected on May 7, 8, 11, 16 and 23, respectively. On May 23, the larva
entered the soil.
D (one larva): One exuvia each was collected on May 7, 8, 11, 15 and 23, respectively. On May 23, the larva
entered the soil.
E (one larva): One exuvia each was collected on May 7, 8, 12, 16 and 23, respectively. On May 23, the larva
entered the soil.
F (16 larvae): Exuviae were not collected. Five larvae (prepupae) went into the soil on May 24, eight larvae on
May 25, one larva on May 26, and two larvae on May 28.
These 21 individuals all died during the hibernation.
On June 1, 2012, Saito found many late-instar larvae staying on the trunk of the Stewartia trees in Mino.
In Wami (230 m alt.), Nakagawa Town, Tochigi Prefecture, Ibuki collected seven larvae (10–18 mm long) on May 23, 2011, but all died by May 29. On June 10 of the same year, five larvae were collected and they went into the soil on June 11. From this series, two male adults emerged on April 16, 2012. On May 4, 2012, Ibuki collected five larvae, four of which grew to the length of about 20 mm by May 22. On May 25, they became prepupae and entered the soil. No adults emerged in 2013.
The egg is inserted into the tissue on the underside of a leaf in its median part or near (but not on) the margin ( Fig. 1 View FIGURE 1 J), usually one egg per leaf. The larva is solitary and does not leave the leaf even if it is disturbed. The prepupa is quite active before entering the soil.
In Kanuma City, the larvae of this species heavily infested the foliage of the host plant almost skeletonizing some of the host trees in 2007 and 2008, but the infestation was not conspicuous thereafter.
Remarks. In the keys to the world species by Wei et al. (2011), Wei & Zhou (2012) and Wei & Niu (2013), this new species does not agree with either of the first couplets. Because of its small size (length 7.5–9.5 mm) and comparatively shallow ventral incision of the clypeus (depth 0.36–0.50 times the entire clypeal length), E. vernalis may agree with the first part of the couplet leading to E. lii Wei, 2011 , from Zhejiang, China, whereas in the entirely pale ventral parts of the abdominal terga and basal ovipositor sheath, the long tarsomere 1 (about 0.82–0.91 times the lengths of the tarsomeres 2–5 together) and the short malar space (about 0.4–0.6 times the diameter of the middle ocellus), E. vernalis does not agree with E. lii but it better agrees with the second part of the couplet.
From the other congeners from Japan, E. vernalis is easily separated by the dark color pattern and the comparatively shallow ventral incision of the clypeus, as noted in the key below. For the moment, we are not able to distinguish the larva of E. vernalis from that of E. nigromaculata , which feeds on the same plant.



| Hyogo |
Museum of Nature and Human Activities |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |