
Dopasia sp.
|
publication ID |
https://doi.org/ 10.5252/g2011n2a10 |
|
persistent identifier |
https://treatment.plazi.org/id/038387F3-FFD7-DE1C-381D-FAC6B55CF941 |
|
treatment provided by |
Marcus |
|
scientific name |
Dopasia sp. |
| status |
|
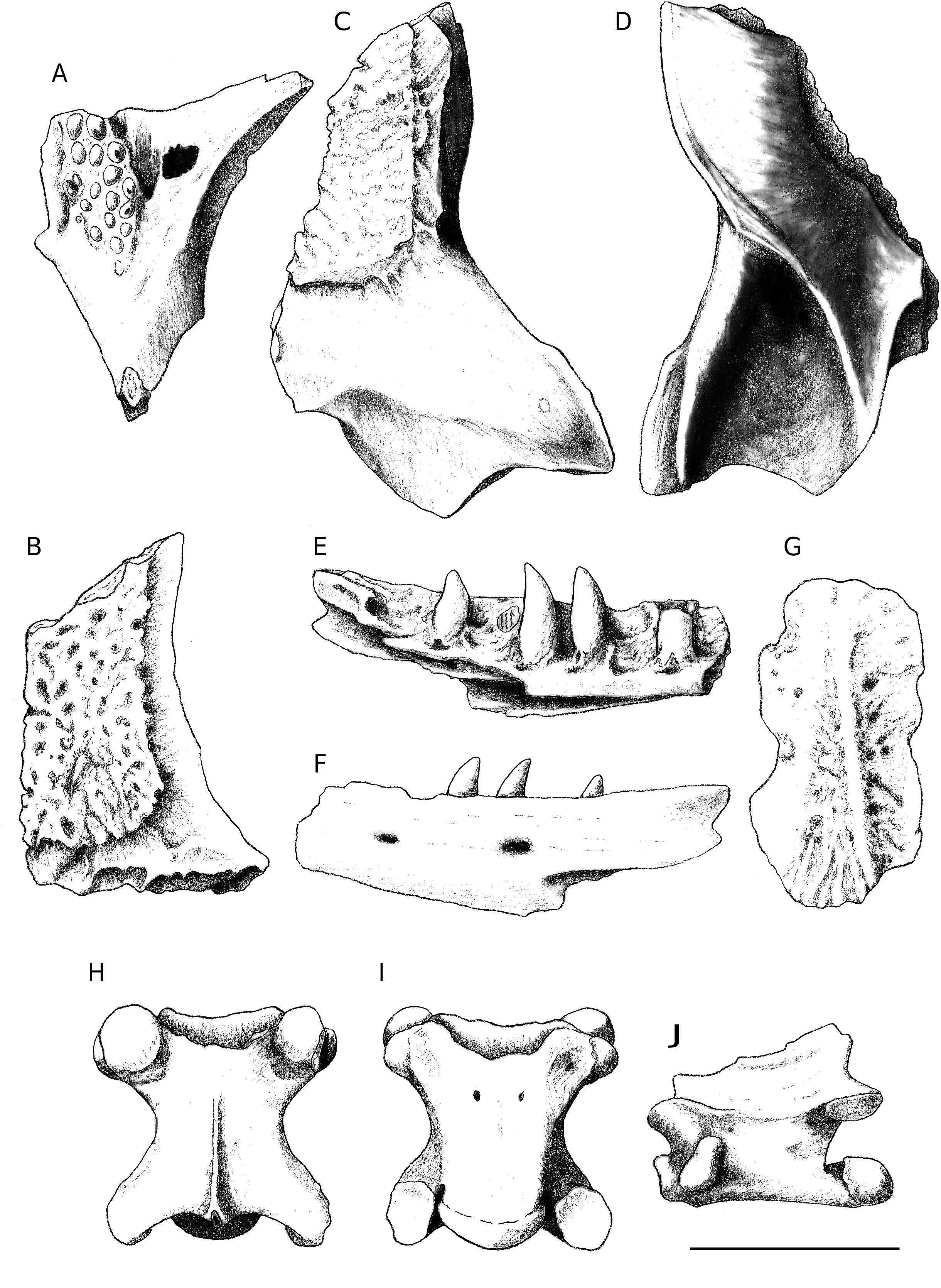
Dopasia sp. ( Fig. 5 View FIG )
MATERIAL EXAMINED. — Maxilla: 2; pterygoid: 2; frontal: 2; parietal: 1; dentary: 1; cervical vertebra: 1; trunk vertebra: 30; caudal vertebra: 33; rib: 11; osteoderm: 5.
DESCRIPTION
The two fragmentary maxillae (an anterior and a posterior fragment belonging to one or two left maxillae) are characterized by canine-like teeth that are slightly curved and have a narrow base. On one of the posterior teeth, a weak ridge is developed along the anterior edge of the apex.
The two pterygoid fragments correspond to the central area of the bone only. The palatine, transverse, and posterior processes are broken off. In ventral view, the pterygoid fragments are characterized by teeth arranged in three or four rows placed in an oval dentigerous region. The 17 or 18 teeth are weakly developed (in some cases there is just a hint of a tooth) and have blunt apexes. A foramen is visible at the level of the pterygoid sulcus in one of the pterygoids.
Both frontals do not preserve the anterior region. They show an evident dermal vermicular sculpture on the dorsal surface; the sculpture does not cover the entire surface of the bone and its lateral edge is marked by a distinct step. In lateral view, the frontals have, in their anterior sector, a deep and well-delimited V-shaped groove whose rims open in anterior direction (= prefrontal impression). Posteriorly, the impression of the postfrontal is visible along the parietal-postfrontal process. The ventral frontal process is moderately prominent and has a convex ventral edge.
The fragmentary parietal is about 5 mm long and it corresponds to the right half of the element whose anterior tip and parotic process (= processus supratemporalis sensu Klembara 1979) are broken off. In dorsal view, this element shows a vermicular dermal ossification, the so-called sculpture, corresponding to the parietal scale; laterally, such dermal ossification develops a subtle dermal “blade” that overhangs the lateral edge, which is markedly convex and corresponds to the anterior branch of the parietal crest. The preserved portion of the parotic process has an undulated surface (clearly visible along the section): convex medially and concave laterally. In ventral view, the anterior branch of the parietal crest is distinctly convex and constitutes the lateral edge of the bone. Posteriorly, the crest becomes concave (= internal branch of the parietatal crest, sensu Fejérváry-Lángh 1932) and it is prolonged up to the medial margin of the parotic process. The junction of this crest with the “ margo posterior of lamina parietalis ” (sensu Klembara 1986) constitutes a concave surface whose posterior extremity extends beyond the posterior rim of the parietal fossa (that occupies about half of the total length of the parietal notch). The posterior branch of the parietal crest has a rounded section; it is more massive anteriorly, where it constitutes the lateral edge of the parotic process, and then becomes slightly thinner and runs parallel to the lateral edge.
A fragmentary left dentary is represented by approximately the posterior two thirds of the bone, likely having eight tooth positions. It is small (the fragment is about 4 mm long) and elongated (maximum height of 1.2 mm). The preserved teeth are subpleurodont, with a narrow base, canine-like and weakly hook-like, with pointed and posteromedially directed apexes; they exceed the height of the lateral wall of the dentary of about one third of their height. In medial view, the Meckel’s canal is partially visible posteriorly but not anteriorly, where it runs along the ventral edge of the bone and is therefore hidden by the horizontal lamina. The latter is inclined medioventrally, except at the level of the last two tooth positions where its remnants are nearly horizontal. The dental groove (sulcus dentalis sensu Fejérváry-Lángh 1923) is underdeveloped. The splenial spine is located under the fourth last tooth position. The splenial crest of the horizontal lamina is distinctly notched between the fourth last and the penultimate tooth positions; this notch represents the dorsal edge of the anterior inferior alveolar foramen. The posterior area of the medial edge of the horizontal blade (carina coronoidea sensu Fejérváry-Lángh 1923) apparently shows a process that might possibly be the result of a breakage or that might correspond to the anterior limit of the insertion area of the coronoid. The coronoid process is moderately robust and posteriorly truncated (it is not unlikely that it is apically broken); its medial surface is slightly overhanging. The coronoid process is approximately as long as the last dentary socket. The intramandibular septum is partially free ventrally. The interalveolar foramen opens under the penultimate tooth position. In labial view, the dental crest is slightly convex and posterodorsally inclined; the coronoid process is apically blunt. The angular process is only slightly shorter than the coronoid process. The coronoid notch is not symmetric and corresponds to a triangular convexity of the lateral wall of the dentary representing the lateral facet for the attachment of the coronoid. The surangular notch reaches, anteriorly, the level of the third last tooth position. The intramandibular septum, as well as the angular process, are partially broken off.
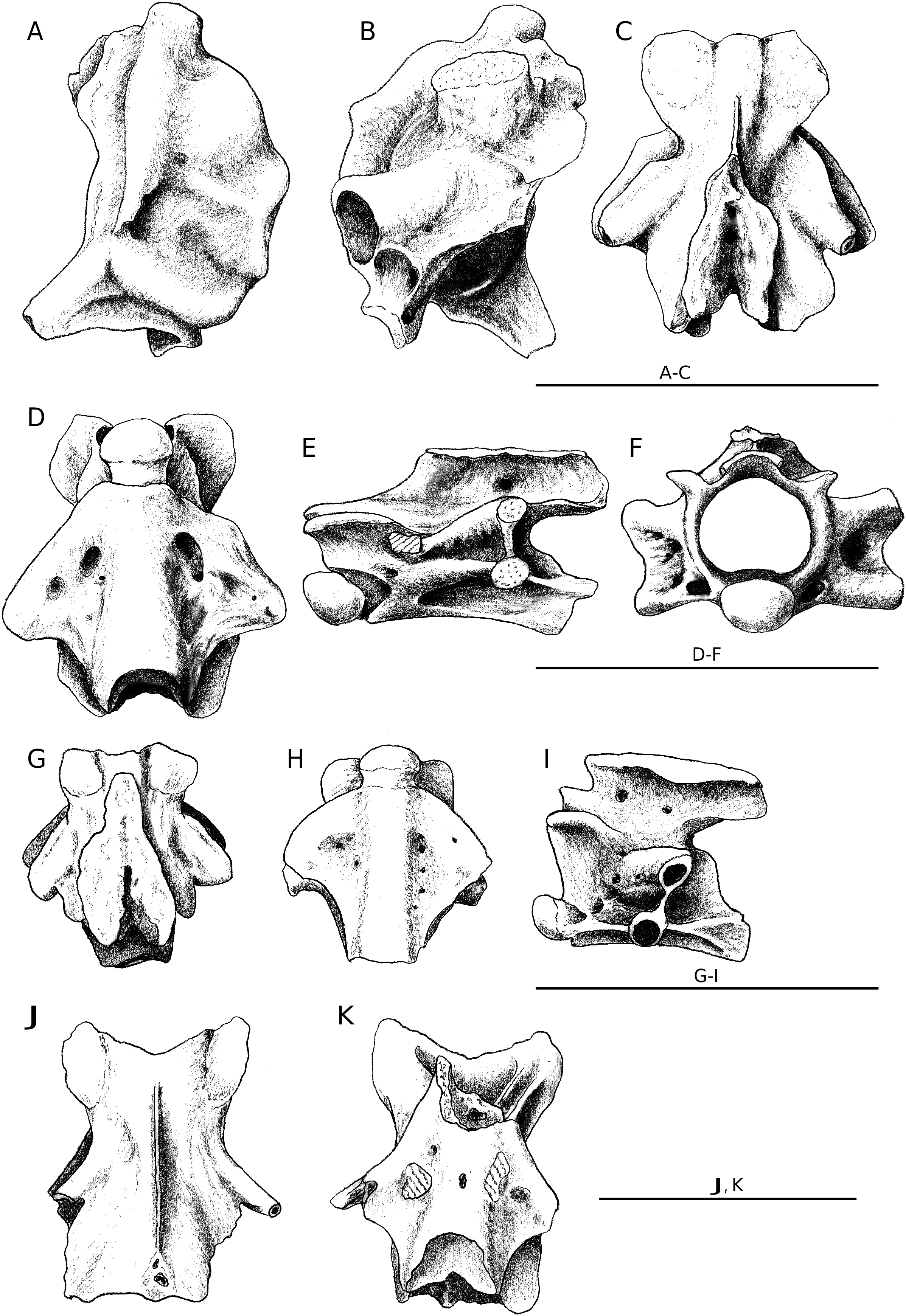
Trunk vertebrae are small, having a centrum length not exceeding 3 mm (but the best-preserved vertebrae are much smaller). The cotyles and condyles are oval in shape; centra have a distinctly flattened ventral surface, with lateral edges anteriorly divergent, slightly concave, and with parallel sides only in the more posterior area; two distinct foramina open on the ventral surface posteriorly to the cotyle; synapophyses are oval-shaped and slightly inclined; prezygapohyses are dorsally tilted, rather large and nearly circular in shape; a laminar neural crest was present but it is invariably broken; the anterior edge of the neural arch is broken but it seems that it was convex; the posterior edge of the neural arch has a neural spine that does not extend beyond the postzygapophyses.
All the caudal vertebrae, except the best-preserved one, are represented by the posterior part of the vertebra only. The best-preserved element has a centrum length of about 1.5 mm but the size of the fragments indicates that some of the vertebrae were significantly larger. Common characters are the oval shape of condyles and cotyles; the flattening of the ventral surface of the centrum; the presence of flattened lateral processes in the anterior region of the vertebra, and of an underdeveloped neural spine that gives origin to a medial process in correspondence of the posterior edge of the neural arch; the development of two parasagittal shallow grooves in the cranial sector of the ventral surface of the centrum; the presence of haemal processes fused to the centrum; the presence of an autotomy surface that is testified by the symmetry of the breakage and its regular surface.
The ribs show an approximately rounded articular surface, a distinct tuberculiform process, and a blade-like ridge in the proximal area.
The osteoderms are lightly built and rather small: the largest is 3.2 mm long and 1.5 mm wide, but one of the fragments, 2.8 mm wide, indicates that the size could have been sensibly larger. All the osteoderms are characterized by an anterior smooth transversal strip followed by an ornate, vermicular, surface. A smooth sagittal keel is present in all the osteoderms except in the above mentioned large fragment.
DISCUSSION
The fossils described above, show the typical characteristics of Anguidae , and more precisely of Anguinae: the dermal ornamentation not extended on the lateral edges of the frontal; the notch of the anterior inferior alveolar foramen; the ventral edge of the intramandibular septum partially visible in labial view; the ventral position of the Meckel’s canal along the anterior edge of the dentary; the surangular impression more extended than the coronoid; the trunk vertebrae with a flat or slightly convex centrum ( Augé 2005, and literature therein).
Despite some contrasting results ( Sullivan 1987; Sullivan et al. 1999), the monophyly of Anguinae seems to be confirmed from both an osteological ( Augé 2005) and molecular ( Macey et al. 1999) point of view, but the systematics of several taxa within the clade is still debated. On the basis of the parietal and tooth morphology, Klembara (1979, 1981) recognized three valid genera for the European Neogene anguids: Pseudopus Merrem, 1820 (for the taxa, formerly referred to Ophisaurus , with molariform posterior teeth and with the parietal characterized by an anterior branch of the parietal crest medially developed and without a distinct parietal notch), Anguis Linnaeus, 1758 , and Ophisaurus Daudin, 1803 (including the taxa with canine-like teeth, an anterior branch of the parietal crest laterally developed and with a distinct parietal notch). According to Gauthier (1982), Sullivan (1987), and Augé (1992), two groups should be identified within the genus Ophisaurus ; the American taxa (with teeth approximately cylindrical and with apexes finely striated and slightly pointed) should be distinguished from the Euro- Asiatic taxa (characterized by canine-like teeth, with non-striated curved apexes). More recently, Augé (2005) suggested to refer the Neogene and Quaternary European anguids to the genera Anguis , Dopasia and Pseudopus mostly on the basis of the morphology and number of teeth, as well as the morphology of the vertebrae and of the osteoderms. Dopasia groups the old world species previously placed in Ophisaurus s.l., and therefore the name Ophisaurus (s.s.) should be restricted to the American representatives of Ophisaurus s.l. Noteworthy is that among the African and Asian taxa with canine-like teeth, only Dopasia harti (Boulenger, 1899) has been examined by Augé and that therefore the referral of the Old World species to the genus Dopasia should be confirmed by further studies; moreover, the genetic differences (mtDNA) observed by Macey et al. (1999) for the African D. koellikeri Günther, 1873 could suggest the revalidation of the name Hyalosaurus koellikeri and to consider the group Ophisaurus (including the American and Asian taxa) as a metataxon whose precise relationships are still to be defined. However, the nomenclature proposed by Augé (2005) has been adopted here.
If compared with extant genera, the fossils from Capo Mannu clearly match the morphology of the genus Dopasia only. Pseudopus is characterized by a medial position of the anterior branch of the parietal crest and by a well-developed lateral facies muscularis (sensu Féjerváry-Lángh 1923); the parietal notch is absent or weakly developed; the teeth are more numerous and the posterior ones are molariform in shape; moreover all the bones are large in size and robust ( Klembara 1979, 1981). In Anguis , whose size is comparable to that of the fossils from Capo Mannu, the number of teeth in a similar portion of dentary is lower; the tooth morphology is more distinctly hooked; the notch of the anterior inferior alveolar foramen is located below the last two tooth positions; the ventral surface of the parietal is not intersected significantly by the arched keel, and, consequently, the parietal fossa is reduced and the parietal notch is more developed; the pterygoids are toothless; the lateral edges of the trunk vertebrae are less divergent; the osteoderms are rounded and devoid of any keel.
The parietal from Capo Mannu differs from the Early Miocene taxa from Dolnice (see Klembara 1979, 1981) D. spinari ( Klembara, 1979) and D. fejfari ( Klembara, 1979) (according to Estes [1983], the morphology of the parietal, that is to say the element used for the diagnosis of these two species, does not allow to discriminate them from other anguines and therefore they should be considered as nomina dubia). Moreover, it differs from extinct D. (? Anguis ) robustus ( Klembara 1979) and extant D. harti , D. koellikeri (the morphology of the latter has been evaluated on the basis of the material coming from a Neolithic site in Morocco, see Stoetzel et al. 2008), and Anguis fragilis Linnaeus, 1758 . The parietal crest has an anterior branch more convex and laterally placed and an inner branch slightly more concave than in Anguis . In dorsal view, the lateral edge (the dorsal surface of the anterior branch of the parietal crest) of the fossil fragment is well visible and convex. The median narrowing is more marked and more anteriorly placed than in the other taxa. Moreover, in D. spinari , D. harti and D. koellikeri , the parotic process is less divergent (this morphology resembles slightly that of D. fejfari , D. [? Anguis ] robustus, and Anguis fragilis ) and the posterior branch of the parietal crest is located on the lateral edge of the parotic process.
Following Augé (2005), the presence of several rows of teeth on the pterygoid is a character restricted to the American taxa, therefore Ophisaurus , whereas in Dopasia and Pseudopus only one row should be present (character 27-0; Augé 2005). However, this character seems to be highly variable and, for the time being, should not be taken into account because several rows are present in all the extant P. apodus at our disposal (5 specimens) and in P. cf. P. pannonicus of the Late Pliocene of Montoussé 5, France ( Bailon 1991), or in the pterygoids of the Miocene of Dolnice referred to Ophisaurus by Klembara (1981) and Roček (1984) (= Dopasia sp. , this work).
The hook-like maxillary and dentary teeth (without striated apexes), is a character of the group Anguis - Dopasia ( Augé 2005) . However, the teeth are less in number, more spaced, taller and more hook-like in Anguis than in the fossil from Capo Mannu and in Dopasia .
The general shape of the teeth of the fragmentary lower jaw from Capo Mannu matches that of the material from the Early Miocene of Dolnice (D. cf. D. spinari ; Roček 1984) and Sansan ( Dopasia sp. 1 ; Augé & Rage 2000) and from the Late Miocene of Polgardi 4 and 5 ( Dopasia sp. ; Venczel 2006). In aff. “ Ophisaurus ” from the Pliocene of the Iles Medas ( Bailon 1991), teeth are more cylindrical and more numerous (thus somehow recalling the American taxa; a general revision of this material is required to address this issue), whereas in D. harti the teeth are more robust, with a basal portion much more enlarged.
While analyzing fossil anguids from Central Europe, Böhme (2002) indicated that sharp anterior cutting edges occur on teeth of maxillae and dentaries associated with D. fejfari parietals, whereas teeth associated with D. spinari parietals do not have such a sharp cutting edge; consequently, the author identified the material from the Middle Miocene from Obergänserndorf ( Austria) as D. aff. D. fejfari . The same taxon (identified as D. cf. D. fejfari ) has been identified in the Early Miocene of Oschiri (Sardinia; Venczel & Sanchiz 2006). In this respect, it is worth mentioning that a keel is present on a tooth of one of the fragmentary maxillae from Capo Mannu, it is nonetheless much less developed than that shown by D. aff. D. fejfari ( Böhme 2002: pl. I, fig. O) or D. cf. D. spinari (Roček 1984: pl. VIII, figs 4, 5; pl. IX, figs 2, 3). The variability of this morphology should be carefully evaluated in order to properly assess its diagnostic value for the identification of anguines.
The morphology of the fossil dentary from Capo Mannu matches well the slender dentaries referred to D. cf. D. spinari (Early Miocene, Dolnice; Roček 1984) and Dopasia sp. 1 (Early Miocene, Sansan; Augé & Rage 2000) and differs from Dopasia sp. from the Late Miocene of Polgardi 4 and 5 which is comparatively more robust (Venczel 2006; Fig. 2A, B View FIG ). Conversely, the fragmentary parietal from Capo Mannu differs from the fossil and extant anguines we used for comparisons, possibly indicating a different specific status. However, the scarcity of the fossil material as well as the imprecise knowledge of the variability of some of the morphological characters of extant and fossil anguine taxa, does not allow any specific identification and the material from Capo Mannu is referred to Dopasia sp.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.