
Dondice trainitoi, Furfaro & Mariottini, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4731.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:25C83363-448C-45E7-8679-FB878E5170B1 |
|
DOI |
https://doi.org/10.5281/zenodo.5919398 |
|
persistent identifier |
https://treatment.plazi.org/id/5072B304-A2F2-4AB0-A0EF-6DF26EC0242B |
|
taxon LSID |
lsid:zoobank.org:act:5072B304-A2F2-4AB0-A0EF-6DF26EC0242B |
|
treatment provided by |
Plazi |
|
scientific name |
Dondice trainitoi |
| status |
sp. nov. |
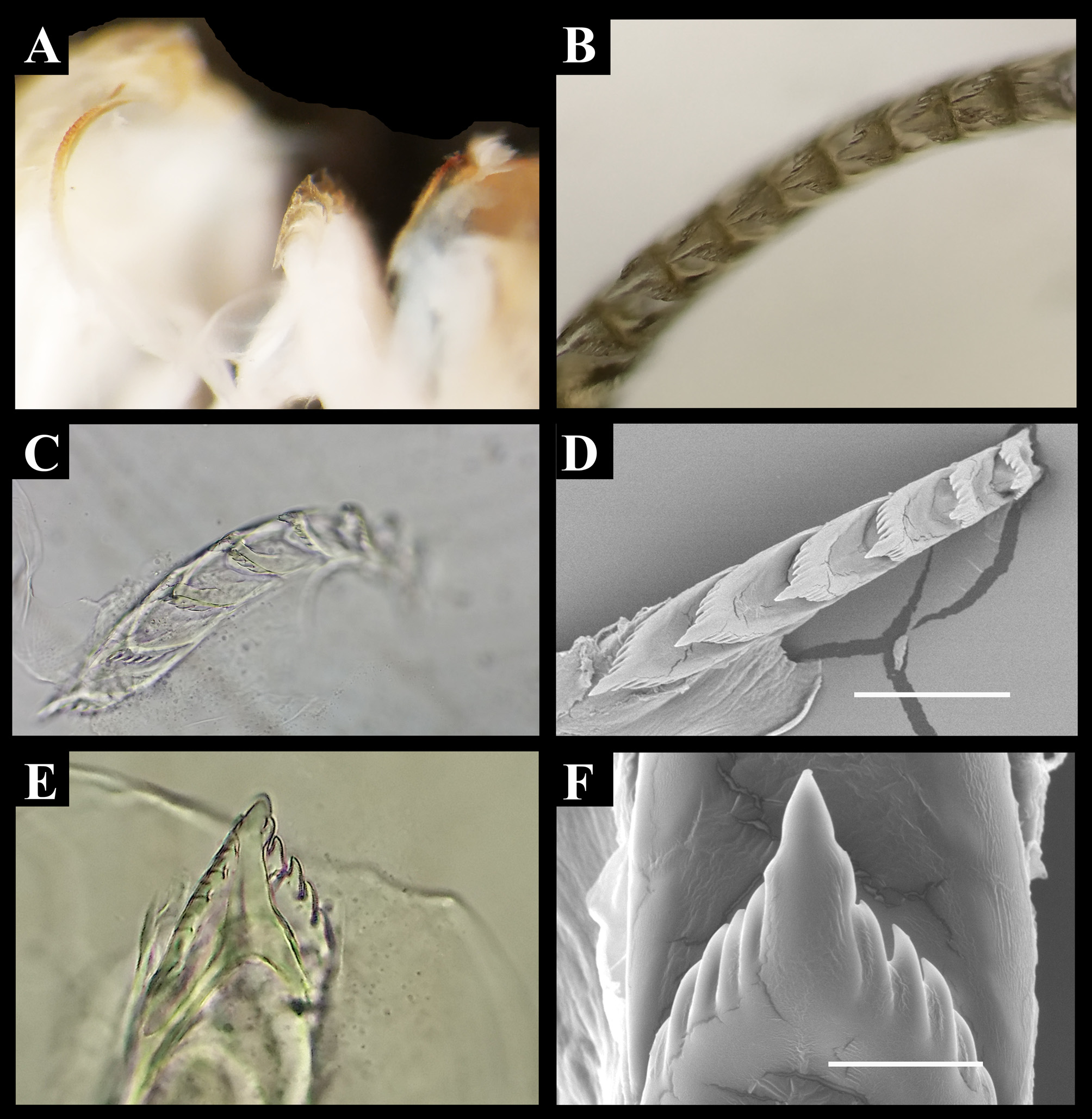
Dondice trainitoi sp. nov. ( Figures 1 A - F View FIGURE 1 )
Zoobank: urn:lsid:zoobank.org:act:5072B304-A2F2-4AB0-A0EF-6DF26EC0242B
Holotype: MNHN IM-2000-33722 , 10 mm in length, Riva di Traiano , Civitavecchia, Latium, Italy, Mediterranean Sea, August 03 2017, 28 m depth, 42°03’15’’N, 11°47’45’’E. GoogleMaps
Paratypes: paratype MNHN IM-2000-33723 , 5 mm in length, Riva di Traiano , Civitavecchia, Latium, Italy, Mediterranean Sea, June 11 2016, 30 m depth; paratype MNHN IM-2000-33724 , 5 mm long, Riva di Traiano , Civitavecchia, Latium, Italy, Mediterranean, June 04 2016, 32 m depth; paratype RM3 _1101, 8 mm in length; paratype RM3 _1102, 11 mm in length, Riva di Traiano , Civitavecchia, Latium, Italy, Mediterranean Sea, August 03 2017, 30 m depth; paratype RM3 _621, 16 mm in length, Riva di Traiano , Civitavecchia, Latium, Italy, Mediterranean Sea, August 25 2016, 30 m depth. All type material sampled at the “Asia” wreck on hydroids .
Etymology: The species name is after Egidio Trainito, expert in nudibranch biology and Mediterranean marine biodiversity, underwater photographer and good friend of the authors.
Holotype morphological description: The body is slender, with a narrow foot. The anterior part of the foot is bilabiate and extended into well-defined propodial tentacles. The elongate body is translucent cream, with bright white spots along the border of the foot, oral foot corners and tail. A characteristic iridescent light blue band is present on the tail, along the dorsum and the head where it bifurcates into two lines ending at the half part of the oral tentacles. Two other lateral lines of the same iridescent color are present dorsally, starting from the lateral part of the head below the rhinophores, becoming narrower and lighter through the body and terminating in the tail joining the dorsal band. A typical black pigment covering the mandibles is clearly visible through the epithelium. The cerata are translucent cream with a digestive gland that is yellowish in the basal portion but becoming gradually orange in the sub terminal part. The upper part of the cerata has a brightly yellowish ring larger in the anterior part than in the back where it is tapered. At the base of the rhinophores, the head is diaphanous, allowing the dark eyes to be seen ( Figs 1A View FIGURE 1 , C-E).
The rhinophores are lamellate with ten dish-shaped annulations and end with a cylindrical tip ( Fig. 1D View FIGURE 1 ). The basal portion and the narrow posterior furrow of the rhinophores are smooth. In their upper portion, there is a brightly white/yellowish stripe that is wider in the anterior than in the posterior part. The oral tentacles are longer than the rhinophores, cylindrical and slender with a bright band in the same color of the apical portion of the rhinophores. The notum showed four clusters of cerata composed by two rows, with the exception of the last group that has only one row of small cerata. There is one precardiac cluster with 22 cerata, two median groups with 16 and 14 cerata respectively, and the posterior one with a single row of 5 cerata. The cerata are slender and cylindrical, light orange, and with a yellow ring which is expanded in the front part and jointed in the rear ( Fig. 1 View FIGURE 1 ). The head is slender. The anus is cleioproct, located within the first post-hepatic arch.
Internal anatomy: Three specimens, including the paratype MNHN IM- 2000-33724 ( Fig. 1D View FIGURE 1 ), were examined. The masticatory jaws are covered with a typical black epithelium ( Fig. 1E View FIGURE 1 ). The underlying chitinous jaws are visible from the muscular lips ( Fig. 3A View FIGURE 3 ). These structures are pale yellowish and characterized by 16 well-developed, triangular denticles per each side ( Figs 3 View FIGURE 3 B-D). The radular formula is 8-12 x (0.1.0) ( Figs 4 View FIGURE 4 A-D). The rachidian tooth has a median cusp, bearing one denticle for each side, and it has 4-5 triangular denticles on each side of the cusp ( Figs 4E, F View FIGURE 4 ). The reproductive system ( Fig. 5 View FIGURE 5 ) is diaulic. The genital openings are on the right side of the body, between the first and the second group of cerata ( Fig. 5B View FIGURE 5 ). The long tubular ampulla runs the length of the female gland mass and the hermaphroditic duct then bifurcates into the oviduct and the vas deferens ( Figs 5 View FIGURE 5 D-F). The male gland terminates into the globular and unarmed penis ( Fig. 5F View FIGURE 5 ). The penial bulb is smooth and muscular externally. The vagina is connected to an ovoidal receptaculum seminis and to the convoluted duct of the female gland mass. The female gland mass is white and complex, with a central rounded and a lateral tortuous mass ( Fig. 5F View FIGURE 5 ).
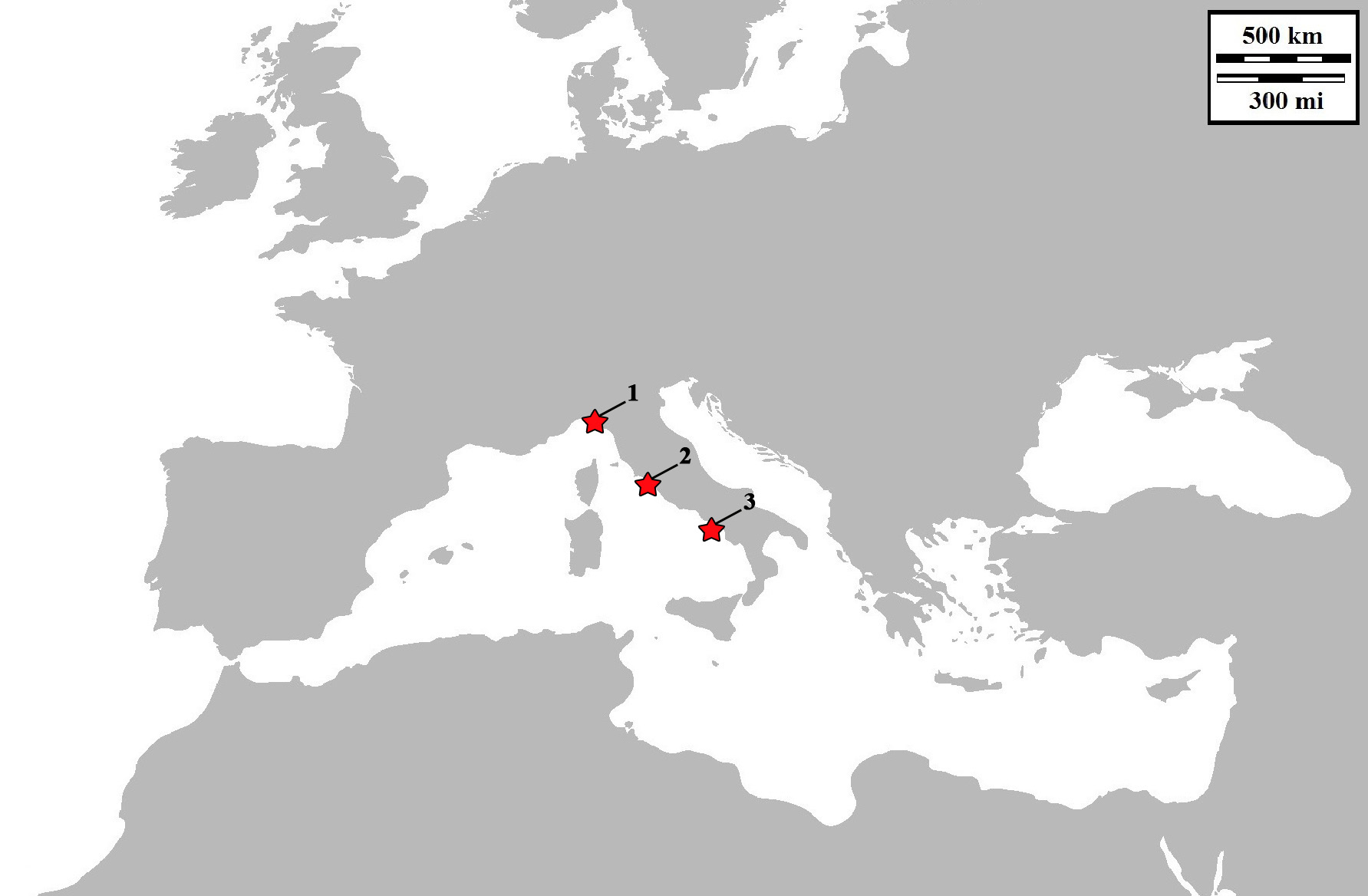
Distribution: Currently, this species is known from the Latium coast (Civitavecchia), type locality, and from Portofino Marine Protected Area (MPA) (Gulf of Genoa, Liguria) and Procida island (Naples, Campania) which are to date respectively the northernmost and the southernmost distribution sites. In particular, most of the findings took place on the ‘Asia’ and the near ‘Liburna’ wrecks, located near the harbour of Civitavecchia where this sea slug is really abundant (with a total of 15-20 individuals observed per each dive). Four years after the first finding in Civitavecchia, about seven individuals were observed from Portofino MPA, while only two specimens were photographed and collected from Procida Island. The specimens were found feeding on colonies of hydroids covering the artificial substrate, made of torn nets from fishing trawlers, wrapping both wrecks, or on colonies of hydroids in a Coralligenous rocky bottom, from June to early September. The geographic distributions of the species is reported in Figure 6 View FIGURE 6 .
Remarks: The morphological variability shown by living animals ( Fig. 1 View FIGURE 1 ) consisted in the body length of the specimens (ranging from 8 to 15 mm), rhinophores with usually 10 or 12 (range 6-17) dish-shaped annulations and the notum with 4 or 5 clusters of cerata. The precardiac cluster of cerata could have up to 24 cerata, the two median clusters up to 18 cerata each one and the posterior one with a single row of maximum 6 cerata. The new species shows 15% mean p -distance at the COI with D. occidentalis and 18% with the Mediterranean D. banyulensis . It also differs morphologically from the three formerly known congeners (Table 3). Dondice trainitoi sp. nov. can be distinguished from D. banyulensis by the presence of the black pigment covering the jaws and forming two black patches clearly visible at the cephalic portion. The only other species which shares this very typical character is the D. occidentalis ‘ sensu Marcus Er. ’ (1958), who described a specimen from Cananèia (San Paolo, Brazil) which ‘ …differs from the Jamaican specimens in some details. The black pigmented epithelium covering the jaws of our species is very striking, even in living slugs, but apparently is wanting in the Jamaican material as Engel does not mention it. ’ (Marcus Er. 1958). Dondice occidentalis ‘ sensu Marcus’ however, has ‘…a median red stripe along the head and a stripe along each side ’ (see the Brazilian specimen photographed by Lindner A. 2014, page 48) that clearly differs from the white ones characterizing D. trainitoi sp. nov. Dondice galaxiana is morphologically entirely different from D. trainitoi sp. nov., in fact it has rhinophores with large dish-shaped annulations, the cerata are clustered on raised cushions and its body has rhomboid-shaped patches on the dorsum between the cerata. These features are completely lacking in the new species. Dondice parguerensis is morphologically similar to D. occidentalis but differing from it for its particular diet which consists of Cassiopea sp. and by the epithelium covering the jaws that is brown amber ( Brandon & Cutress 1985) and not dark or black as in D. occidentalis .
Specimens of the new species were observed and photographed in situ feeding on a reddish/orange hydroid belonging to the family Campanulariidae Johnston, 1836 . The egg mass is a white lace coiled around the hydroid ( Fig. 1F View FIGURE 1 ). Interestingly, as known for other facelinid species, D. trainitoi sp. nov., when disturbed, autotomized its cerata to distract potential predators.



| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |