
Dipolydora spinosa, Radashevsky, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5162.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:11BA9195-9A8E-4098-975B-E010627F9DFD |
|
DOI |
https://doi.org/10.5281/zenodo.6817797 |
|
persistent identifier |
https://treatment.plazi.org/id/CE6B878F-E52F-4B92-866F-456FF0FEA34A |
|
taxon LSID |
lsid:zoobank.org:act:CE6B878F-E52F-4B92-866F-456FF0FEA34A |
|
treatment provided by |
Plazi |
|
scientific name |
Dipolydora spinosa |
| status |
sp. nov. |
Dipolydora spinosa View in CoL sp. nov.
http://zoobank.org:act: CE6B878F-E52F-4B92-866F-456FF0FEA34A
( Figs 4−10 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
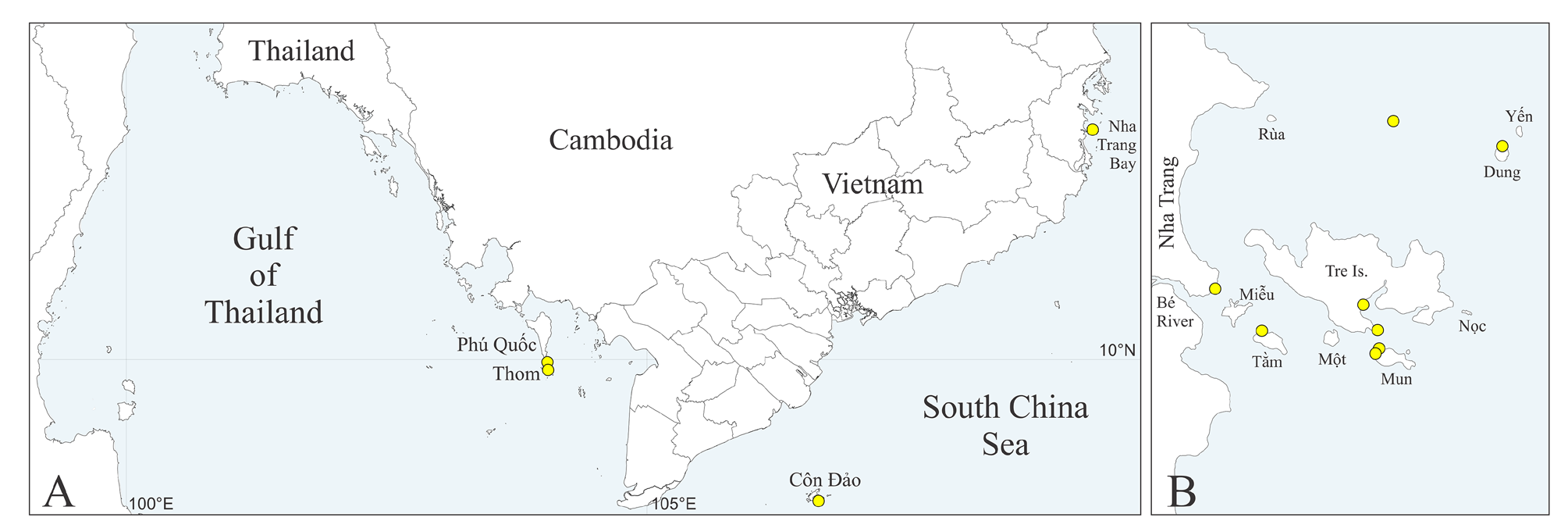
Type material. South-China Sea, Vietnam, Gulf of Thailand, Phu Quoc Is., 0.5 m, in shell of saddle oyster Placuna ephippium (Philipsson) , 3 Feb 1986, MIMB 42712 (paratype). Côn Dao Is., 5 m, in shell of the oyster Lopha cristagalli (Linnaeus) , 19 Feb 1986, MIMB 42713 (paratype). Nha Trang Bay: Dung Is., 10 m, in shells of barnacle, Chama sp. and Millepora dichotoma Forsskål , 2 Jun 2006, MIMB 42714 (8 paratypes); Mun Is., 10 m, in hump coral Porites lutea Milne Edwards & Haime , 7 Jun 2006, 42715 (paratype); Dung Is., 10 m, in empty shell of the oyster Lopha cristagalli , 10 Jun 2006, 42716 (paratype); Dung Is., 10 m, in hump coral Porites lutea , 20 Jun 2006, 42717 (18 paratypes); Mun Is., 10 m, in hump coral Porites lutea , 30 Jun 2006, 42718 (21 paratypes); Tre Is., 5 m, in shell of saddle oyster Placuna ephippium , 26 Apr 2021, 42719 (2 paratypes); Tre Is., 3 m, in dead coral encrusted by coralline alga, 30 Apr 2021, 42720 (2 paratypes), 42721 (holotype) . Complete data on material examined is provided in Supplementary Table S2.
Adult morphology. Adults up to 35 mm long, 1 mm wide for 140 chaetigers. Holotype originally 25 mm long, 0.9 mm wide for 107 chaetigers; anterior and posterior parts fixed in formalin; 35 middle chaetigers fixed in ethanol for molecular analysis. Body light tan in life. Juveniles to about 70-chaetiger stage with remains of larval pigmentation on 15−22 anterior chaetigers, comprising paired dorsal and dorso-lateral melanophores, median dorsal vesiculate melanophores from chaetiger 3, lateral melanophores from chaetiger 2, and small paired ventral melanophores from chaetiger 3; black pigment between eyes, and one pair of melanophores in lateral lips of peristomium ( Figs 4A, B View FIGURE 4 , 10D View FIGURE 10 ).
Prostomium anteriorly bifurcated ( Figs 4A View FIGURE 4 , 5A, B View FIGURE 5 ), posteriorly extending to middle of chaetiger 5 as a low caruncle, shorter in small individuals ( Fig. 6A View FIGURE 6 ) (over 1/3 of chaetiger 5 in holotype). Low occipital antenna present on caruncle at level of anterior part of chaetiger 1 ( Fig. 7A View FIGURE 7 ). One to two small black eyes present in small juveniles; large individuals without eyes. Palps as long as 15−20 chaetigers, with frontal longitudinal groove lined with short cilia, latero-frontal motile compound cilia, short compound non-motile cilia arising directly from palp surface and sparsely scattered on lateral and abfrontal palp surfaces.
Chaetiger 1 with capillary chaetae and postchaetal lamellae in both rami; lamellae same as on chaetiger 2; notochaetae as long as but fewer than those on chaetiger 2; neurochaetae shorter than those on chaetiger 2. Juveniles to 30−50-chaetiger stage with only capillary chaetae in posterior notopodia; larger individuals usually with three, occasionally two or four awl-like spines in addition to 1−5 notopodial capillaries in a tuft from chaetigers 28−52 ( Figs 6B View FIGURE 6 , 7 View FIGURE 7 E−H, 8K, L). Spines almost straight, with distal end slightly protruding out of body wall, smooth surface and basal end blunt ( Fig. 8K View FIGURE 8 ).
Chaetiger 5 twice as large as chaetigers 4 and 6, on each side with up to six geniculate dorsal superior capillaries ( Fig. 8F View FIGURE 8 ), six falcate spines alternating with bilimbate-tipped companion chaetae ( Fig. 8 View FIGURE 8 A−D) and arranged in oblique slightly curved row, and six ventral capillaries ( Fig. 8E View FIGURE 8 ). Dorsal superior and ventral capillaries winged, shorter and fewer than those chaetae on chaetigers 4 and 6. Falcate spines with spoon-like hollow on subdistal concave side and narrow transverse shelf on subdistal convex side ( Fig. 8B, D View FIGURE 8 ); transverse shelf apparently worn and therefore not seen on spines of some individuals.
Hooded hooks in neuropodia from chaetiger 7, up to six in a series, accompanied by 1−2 inferior winged capillaries ( Fig. 8G View FIGURE 8 ) until chaetigers 10−12, and alternating with 1−2 hair-like alimbate capillaries ( Fig. 8I View FIGURE 8 ) in 3−10 posterior neuropodia. Hooks bidentate, with slightly curved shaft without constriction ( Fig. 8H, J View FIGURE 8 ).
Branchiae usually from chaetiger 8 (on chaetigers 8−88 in holotype), occasionally from chaetiger 9 (3 of 26 examined paratypes), full-sized from chaetigers 10−11, absent from posterior 1/3−1/4 part of body ( Fig. 6B View FIGURE 6 ), free from notopodial postchaetal lamellae, flattened, with surfaces oriented parallel to body axis.
Nototrochs in juveniles and females from chaetigers 9−13, in males from chaetiger 1, all composed by single rows of ciliated cells. Intersegmental transverse ciliation present on anterior chaetigers in males as single rows of ciliated cells along anterior edge of chaetigers, from chaetiger 2 to chaetigers 10−12.
Pygidium fleshy, cup-shaped with dorsal gap to incision, white due to numerous fusiform glandular cells ( Figs 7 View FIGURE 7 E−G, 9A).
Glandular pouches in neuropodia from chaetiger 6, large from chaetiger 7 to chaetigers 10−12 and then gradually diminishing in size.
Digestive tract with gizzard-like structure beginning from chaetigers 13−29 (from chaetiger 21 in holotype) and extending through 1−2 chaetigers ( Figs 5C View FIGURE 5 , 6A View FIGURE 6 ). Gizzard composed of anterior transparent muscular part and posterior white, apparently secretory, part. Some grain-like structures present in wall of muscular part of some individuals ( Fig. 5D View FIGURE 5 ; these may represent fragmented “teeth” of the gizzard as discussed in Blake 1971, Fig. 14g View FIGURE 14 therein). Rectum white in 5−15 posterior most chaetigers.
Nephridia from chaetiger 7, opening to exterior via two nephridiopores situated on lateral sides in anterior sterile chaetigers and on dorsal side in fertile chaetigers ( Fig. 10E View FIGURE 10 ).
MG staining. Paired blotches (gatherings of strongly stained, probably glandular, cells) appeared on the dorsal side from chaetigers 12–17 onwards ( Fig. 10A, B, E View FIGURE 10 ), except 10−30 posterior most chaetigers. Narrow transverse bands were weakly stained on the ventral side from chaetiger 7 to chaetigers 10–12 ( Fig. 10C View FIGURE 10 ). Prostomium and the peristomium did not absorb the dye, but the foregut was evenly colored ( Fig. 10C, D View FIGURE 10 ).
Habitat. Dipolydora spinosa sp. nov. is an opportunistic borer, making U-shaped burrows in shells of the saddle oyster Placuna ephippium , cockscomb oyster Lopha cristagalli (Linnaeus) , jewel box clam Chama sp. , barnacles, in the hump coral Porites lutea Milne Edwards & Haime , and the net fire coral Millepora dichotoma Forsskål. Up to three worms were found per one square cm of coral surface and hundreds of worms were occasionally present in corals of about 20 cm in diameter.
Reproduction. Dipolydora spinosa sp. nov. is dioecious. Paired gonads are attached to segmental blood vessels from chaetigers 23−42 to chaetigers 50−82 (n=15) ( Fig. 9A View FIGURE 9 ). Smallest mature individuals were about 17 mm long for 70 chaetigers. Holotype is a female with oocytes in chaetigers 28−82. Oogenesis is mainly intraovarian. Developed oocytes accumulate in the coelomic cavity prior to spawning. The coelomic oocytes were subspherical to slightly oval, 100−115 µm in diameter.
In males, spermatogonia proliferate in testes; spermatogenesis occurs in the coelomic cavity. Spermatids are joined in octads ( Fig. 8M View FIGURE 8 ). Spermatozoa are introsperm about 1 µm in diameter, with a pointed acrosome 1.5 µm long, nucleus 7.5 µm, middlepiece 6 µm, head+middlepiece 15 µm long, and flagellum 50 µm long.
A small colony of about 20 individuals in the hump coral Porites sp. collected from Nha Trang Bay in June 2006 contained only males. One of them had seven chaetigers regenerating anteriorly, middle stock chaetigers, and a few chaetigers regenerating posteriorly; regenerating chaetigers were slightly smaller and more transparent than fully developed stock chaetigers. This colony likely appears to have developed due to asexual reproduction by architomy from a single settled larva of male gender. It seems likely that after settlement, individuals first undergo a series of architomic divisions and only after that mature and start reproducing sexually. Such colony formation has been found in other polydorin species that exhibit architomy (Radashevsky, pers. obs.) and will be the subject of future studies.
Larval development. Females lay eggs into transparent capsules which join to each other in a string and each attach by two short stalks to the inner wall of the burrow ( Fig. 9C View FIGURE 9 ). Up to 60 eggs were deposited in one capsule and up to 55 capsules with about 3000 eggs were produced by one female per brood; small females had smaller broods. The laid eggs were 105−120 µm in diameter, with smooth envelope less than 1 µm thick. Females brooding larvae in capsules had the next generation of vitellogenic oocytes developing in the ovaries. Most of the eggs in the broods developed into larvae, but about 10% of eggs were abortive (it is not known whether these result from sperm limitation or represent nurse egg development; see Smith & Gibson 1999). The egg capsules with trochophores and abortive eggs were found in Nha Trang Bay in June 2006 ( Fig. 9C, D View FIGURE 9 ). The trochophores had a few very small trochoblasts bearing short cilia of the prototroch, and one pair of rounded ventral ciliary patches. They had no eyes and moved inside capsules due to active beating of the ciliary patches. The larvae had a small mouth and a short foregut lined with numerous fine cilia ( Fig. 9D View FIGURE 9 ). The posterior end of the foregut was joined to a spherical mass of large yolky endodermal macromeres occupying most of the inner space of larvae. Lumen of midgut, posterior gut, and anus were not yet developed. The duration of larval development inside the capsules remains unknown, but compared to other polydorins producing similar broods (see Blake & Arnofsky 1999), D. spinosa sp. nov. larvae probably hatch after growing 3−5 chaetigers.
Remarks. By having an occipital antenna on the prostomium, D. spinosa sp. nov. is similar to D. tentaculata ( Blake & Kudenov, 1978) from New South Wales, Australia, and D. anatentaculata Delgado-Blas, 2008 from the Gulf of Mexico, Florida. Dipolydora spinosa sp. nov. and D. tentaculata have caruncles of similar maximal length, extending to the middle of chaetiger 5 (for D. anatentaculata, Delgado-Blas 2008: 8 , fig. 2A described the “caruncle extending posteriorly to segment 4” but illustrated it as reaching chaetiger 5). Dipolydora spinosa sp. nov. has a cup-shaped pygidium with only dorsal incision (which is comparatively rare among Dipolydora spp. that mainly have lobate pygidia with three or four lobes) while the pygidial shapes are unknown for D. tentaculata and D. anatentaculata . Dipolydora spinosa sp. nov. and D. anatentaculata are shell-borers while D. tentaculata lives in tubes in sediment. Dipolydora spinosa sp. nov. differs from these two species in the structure of the falcate spines of chaetiger 5. In D. tentaculata and D. anatentaculata , the spines are simple, without accessory structures, whereas in D. spinosa sp. nov. the spines have a spoon-like hollow on the concave side and a narrow transverse shelf on the convex side. Blake & Kudenov (1978: 250) noted that in D. tentaculata the notochaetae of the posterior chaetigers “including long slender capillaries and a stouter spine.” On the illustration ( Blake & Kudenov 1978: fig. 39c), however, the chaeta resembles thick anterior-row capillary notochaeta rather than a genuine spine (see Discussion below). Adults of D. spinosa sp. nov. have 2−4 (usually three) awl-like spines in addition to 1−5 capillaries in a tuft in the posterior notopodia; the spines are almost straight, with basal end blunt.
Etymology. The species name, feminine for Latin spinosus (spiny, thorny), refers to the awl-like spines in the posterior notopodia of adults.
Distribution. South-China Sea, Vietnam (see Table S2, Fig. 1A, B View FIGURE 1 ).








| MIMB |
Museum of the Institute of Marine Biology |
| MG |
Museum of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |