
Diaspidiotus degeneratus (Leonardi), 1896
|
publication ID |
https://doi.org/ 10.5281/zenodo.282977 |
|
publication LSID |
lsid:zoobank.org:pub:68A889B1-51C1-40B2-A264-DE8AE782B94C |
|
DOI |
https://doi.org/10.5281/zenodo.6179512 |
|
persistent identifier |
https://treatment.plazi.org/id/0382231A-FFD2-FFA9-1398-FD60FC1DFB96 |
|
treatment provided by |
Plazi |
|
scientific name |
Diaspidiotus degeneratus (Leonardi), 1896 |
| status |
|
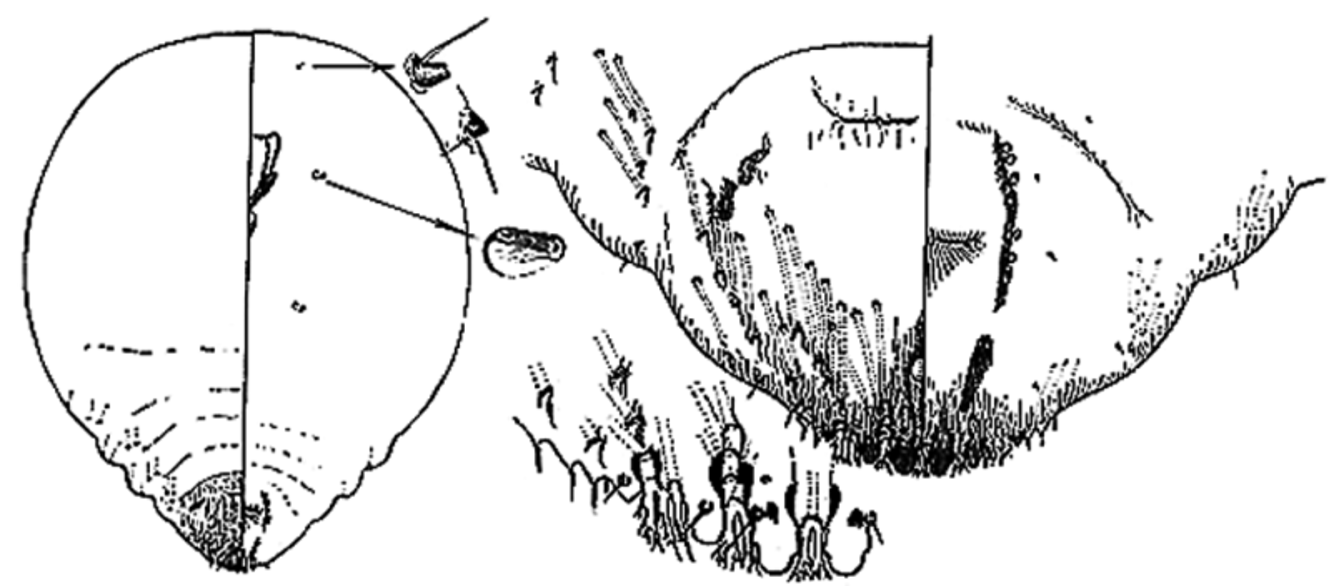
Diaspidiotus degeneratus (Leonardi), 1896 View in CoL ( Fig. 11 View FIGURE 11 ), stat. rev.
Fig. 21 View FIGURE 21
Chrysomphalus degeneratus Leonardi View in CoL in Berlese & Leonardi, 1896: 345. Type data: Italy (Portici) on leaves of Pandanus graminifolia . Type depository: Portici: Dipartimento de Entomologia e Zoologia Agraria di Portici, Universita di Napoli Federico II, Italy
Aspidiotus degeneratus (Leonardi) View in CoL ; Cockerell, 1896: 334.
Aspidiotus (Chrysomphalus) degeneratus (Leonardi) View in CoL ; Cockerell, 1897: 29.
Hemiberlesia degenerata (Leonardi) View in CoL ; McKenzie, 1939: 54.
Abgrallaspis degeneratus (Leonardi) View in CoL ; Balachowsky, 1948: 317; Miller & Davidson, 2005: 42.
Diaspidiotus degeneratus (Leonardi) View in CoL ; Borchsenius, 1950: 225.
Dynaspidiotus degeneratus (Leonardi) View in CoL ; Borchsenius, 1966: 282; Danzig, 1993: 151.
Distribution: Nearctic: USA; Western Palearctic: Georgia, Greece, Italy, Portugal; Eastern Palearctic: China, Japan, North Korea.
Host families: Araliaceae , Aquifoliaceae , Celastraceae , Ericaceae , Oleaceae , Pandanaceae , Rutaceae , and Theaceae .
Discussion: Scale insect workers have found it difficult to place Chrysomphalus degeneratus in an appropriate genus based on its morphology, since it lacks some of the characteristics that define the various genera and/or exhibits characteristics intermediate between genera. Since Leonardi described it in 1896, it has been placed in the following genera: Abgrallaspis , Aspidiotus , Diaspidiotus , Dynaspidiotus and Hemiberlesia . Prior to this study, it was placed in the genus Abgrallaspis . Research on the molecular systematics of diaspidid species by Andersen et al. (2010) and by Rugman-Jones et al. (2010), which included Abgrallaspis degeneratus , Chrysomphalus aonidum , C. dictyospermi , Abgrallaspis cyanophylli (the type species of the genus Abgrallaspis ) and numerous species from several other aspidiotine genera, showed a very close relationship between A. degeneratus and species of Chrysomphalus , Aonidiella and Diaspidiotus and a relatively distant relationship to the Abgrallaspis species included in their study. After discussion with Benjamin Normark (pers. comm.) and consideration of the results of Anderson et al. (2010) and Rugman-Jones et al. (2010) it was decided to examine the morphology of Abgrallaspis degeneratus to determine whether it supported the molecular findings.
Although the genus Abgrallaspis has been shown to be paraphyletic (Andersen et al., 2010; Rugman-Jones et al., 2010) and its limits poorly-defined, it appears that the species currently placed in Abgrallaspis lack the paraphysis in the L2–L3 interlobular space and lack the long and slender paraphysis arising from the mesal corner of L1 typical of Chrysomphalus , Aonidiella and certain species of Diaspidiotus [as well as in A. degeneratus ]. In Abgrallaspis , the paraphysis arising from the mesal corner of L1 is entirely absent, extremely short, or forms a wide basal sclerosis, at least half the width of L1, and the paraphysis arising from the lateral corner of L1 is either absent, or relatively short and/or weakly developed. In addition, most Abgrallaspis species have 3 pairs of well-developed pygidial lobes. Unlike Chrysomphalus species, the paraphyses of A. degeneratus are less well-developed, about as long as, or shorter than, the pygidial lobes, the paraphysis in the interlobular space between L2 and L3 is absent or very short and inconspicuous, and the margin anterior to L3 has a series of 3 short glandular spines, whereas in Chrysomphalus the paraphyses are well-developed, usually longer than the pygidial lobes with a well-developed, elongate paraphysis in the interlobular space between L2 and L3 and the margin anterior to L3 has a series of plates with clubbed, fringed or bifurcate apices. Based on these characters, we believe that A. degeneratus is better placed in Diaspidiotus , the genus in which Borchsenius (1950) had placed it. It is most similar to Diaspidiotus africanus ( Marlatt, 1908) in the presence of perivulvar pores and in the shape of the lobes, paraphyses, plates and spines.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Diaspidiotus degeneratus (Leonardi), 1896
| Smith-Pardo, Allan H., Evans, Gregory A. & Dooley, John W. 2012 |
Dynaspidiotus degeneratus
| Danzig 1993: 151 |
| Borchsenius 1966: 282 |
Diaspidiotus degeneratus
| Borchsenius 1950: 225 |
Abgrallaspis degeneratus
| Miller 2005: 42 |
| Balachowsky 1948: 317 |
Hemiberlesia degenerata
| McKenzie 1939: 54 |
Aspidiotus (Chrysomphalus) degeneratus
| Cockerell 1897: 29 |
Chrysomphalus degeneratus
| Berlese 1896: 345 |
Aspidiotus degeneratus
| Cockerell 1896: 334 |