
Cyclocypris Brady & Norman, 1889
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2820.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03EB87C8-6F6C-FFD2-FF30-FBA5A1CC7746 |
|
treatment provided by |
Felipe |
|
scientific name |
Cyclocypris Brady & Norman, 1889 |
| status |
|
Genus Cyclocypris Brady & Norman, 1889 View in CoL
1889 Cyclocypris Brady & Norman : 70.— Type species: Cypris globosa Sars, 1863 (by monotypy)
1941 Cyclocypria Dobbin : 235.— Type species: Cyclocypria kincaidia Dobbin, 1941 (by monotypy)
1949 Cyclocypris (Leucocypris) Jančařík View in CoL : 10.— Type species: Cypris (Leucocypris) drastichi Jančařík, 1949 (by monotypy).
1995 Cyclocypris (Laevicypris) Krstić View in CoL : 37.— Type species: Cyclocypris laevis (O. F. Müller, 1776) (by original designation).
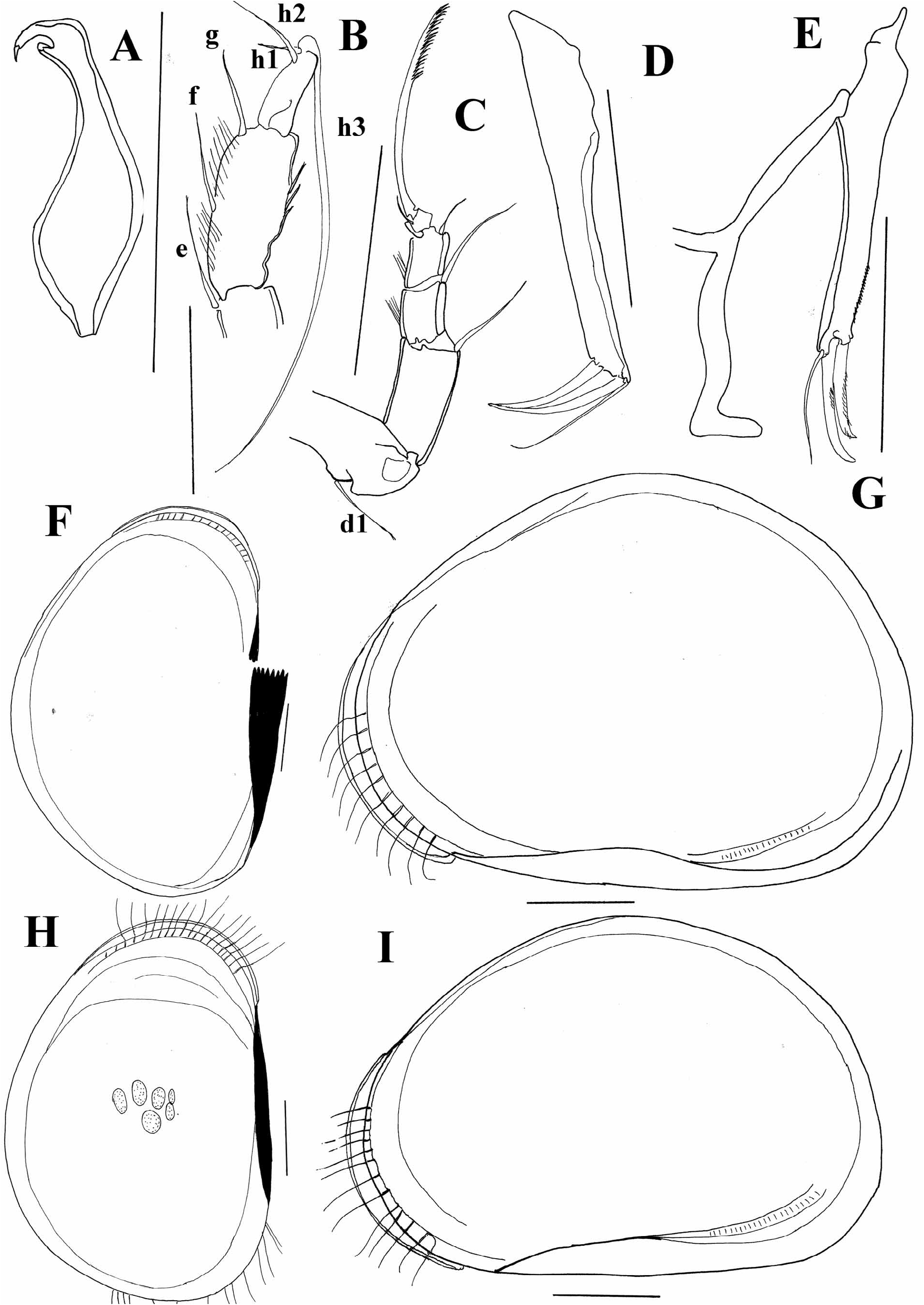
Diagnosis. Carapace short when seen laterally, ovate in dorsal view, with greatest H and W exceeding ½ L. LV overlaps RV both dorsally, frontally and ventrally, or RV overlaps LV. A1 7-segmented. Penultimate segment of A 2 in male subdivided, but t2 and t3 setae not transformed into sexual bristles. Terminal segment of Md palp rather short, L:W ratio being 2:1. Terminal segment of Mxl palp square. Prehensile palps asymmetrical to different degrees. T2 with basal seta, (“d1”) present, same appendage 5-segmented. T3 4-segmented. Basal segment with d1, d2 and d3 setae developed. Penultimate segment not subdivided, “e”, “f”, and “g” setae long. Terminal segment elongated more than 3 times longer than wide. Setae “h1” and “h2” short, “h3” long. Caudal ramus completely developed, hemipenis with lobes “a”, “b”, and “h” developed. Lobe “a” and “b” being short and stubby. Genital field without any extensions. Zenker organ with 7 whorls of spines.
Type species. Cyclocypris globosa (Sars, 1863) View in CoL .
Other species. C. ampla Furtos, 1933 ; C. brevisetosa Bronstein, 1928 ; C. diebeli Absolon, 1973 ; C. globosa (Sars, 1863) View in CoL ; C. forbesi Sharpe, 1897 [ C. washingtoniensis Dobbin, 1941 syn. nov.]; C. laevis (O. F. Müller, 1776) ; C. mediosetosa Meisch, 1987 ; C. modesta (Herrick, 1887) ; C. nahcotta Dobbin, 1941 ; C. ovum ( Jurine, 1820) View in CoL ; C. pusilla Sars, 1895 ; C. scrobiculata Klie, 1936 ; C. serena (Koch, 1838) ; C. sharpei Furtos, 1933 [ C. cruciata Furtos, 1935 syn. nov.]; C. vinyardi Külköylüoğlu, 2008 .
Remarks and affinities. The genus Cyclocypris is most closely related to Kempfcyclocypris gen. nov., with which it shares a similar chaetotaxy of the male A2, T2 and T3, and the appearance of the prehensile palps. Both genera lack the sexual bristles on the second antenna, and they have a basal seta on the T2, a long “g” seta on the T3. On the other hand, they differ in the number of segments on A1 (6-segments in the new genus, 7 in Cyclocypris ), and the appearance of hemipenis, but in both genera lobes are short and stubby. Also, the terminal segment of T3 is much more elongated in Cyclocypris species than in Kempfcyclocypris . From all other genera, Cyclocypris differs in the appearance of hemipenis. Namely lobes “a” and “b” are thin and elongated in all other genera of Cyclocypridinae , except in Kempfcyclocypris . Other differences are the elongated terminal segment on the T 3 in Cyclocypris and long “g” seta on the same appendage, which is very short, if not even absent, in the other genera of the subfamily Cyclocypridinae .
The species Cyclocypria kincaidia Dobbin, 1941 and Cyclocypris wyomingensis Ferguson, 1966 were not included in the key to species of the genus Cyclocypris . The first species was described from Lake Washington ( Dobbin 1941), as the type and the only species of the genus Cyclocypria Dobbin, 1941 . However, the species was described after juvenile specimens, and the differential diagnosis, which concerns the chaetotaxy of T3 is an artefact of specimens being an early instar. This was already noticed by Dobbin-Evenson (1969) who placed the species in the genus Cyclocypris . The original description and drawings of C. wyomingensis simply do not have enough information to give this species a correct phylogenetical position within the genus, and it extremely resembles C. laevis (O. F. Müller, 1776) . Külköylüoĝlu (2008) has examined the type material, but unfortunately, the little additional descriptions he provided (based on the parts he was able to observe), still do not give enough differential characters for the species. It has to be pointed out that what Külköylüoĝlu (2008) observed was quite different from Ferguson’s (1966) description of C. wyomingensis . Dobbin (1941) described C. washingtoniensis , which was re-examined by Külköylüoğlu (2008). The latter author postulated that this might be a synonym of C. forbesi , but he did not formally synonymise the species. After rechecking several slides deposited at the SMNH (see Material Examined) I here formally synonymise C. washingtoniensis with C. forbesi , based, most of all on the characteristic appearance of the prehensile palps ( Figure 1A View FIGURE 1 ). Unfortunately, all the slides are in a very poor condition, so additional drawings could not be provided.
The species C. cruciata Furtos, 1935 , described from Massachusetts ( Furtos 1935), is here synonymised with C. sharpei Furtos, 1933 , described from Ohio. According to Furtos (1935) C. cruciata differs from C. sharpei in the length of carapace and the presence of the small plates, named “platelets” by Matzke-Karasz et al. (2004) along the anterior end of the RV, which are also present in C. diebeli Absolon, 1973 . The appearance of the soft parts of C. cruciata and C. sharpei are extremely similar ( Figure 1B–E View FIGURE 1 ). Both species miss the posterior seta on the UR, have long anterior seta on the ramus, and have the same hemipenis (only in C. sharpei lobe “a” is slightly more curved towards interior side, what can be due to the slide preparation). Külköylüoğlu (2008) was able to see only the left valve from the collection of the Smithsonian Museum of Natural History, where the type material is deposited. After examining the type material of both species including both LV ( Figure 1F, H View FIGURE 1 ) and RV ( Figure 1G, I View FIGURE 1 ), I came to the conclusion that both C. sharpei and C. cruciata have a normal hyaline flange along the margin of both RV and LV. What probably confused Furtos (1935) is the way marginal setae on the RV are curved, giving the appearance of the “divided” hyaline flange ( Figure 1G View FIGURE 1 ). Unfortunately, slides of C. cruciata are in a poor condition and I am unable to provide additional drawings.
Henry (1923) described Cyclocypris tenuissima Henry, 1923 from an unknown locality in New South Wales. The species description is very short and the drawings imprecise, but it is obvious that the species Henry (1923) was dealing with does not belong in to the subfamily Cyclocypridinae . The posterior seta on the UR is in this species positioned very close to the distal margin, a feature not present in the subfamily. Beside, the UR itself is very thin and long. It is more likely that this species belongs to the family Cyprididae .
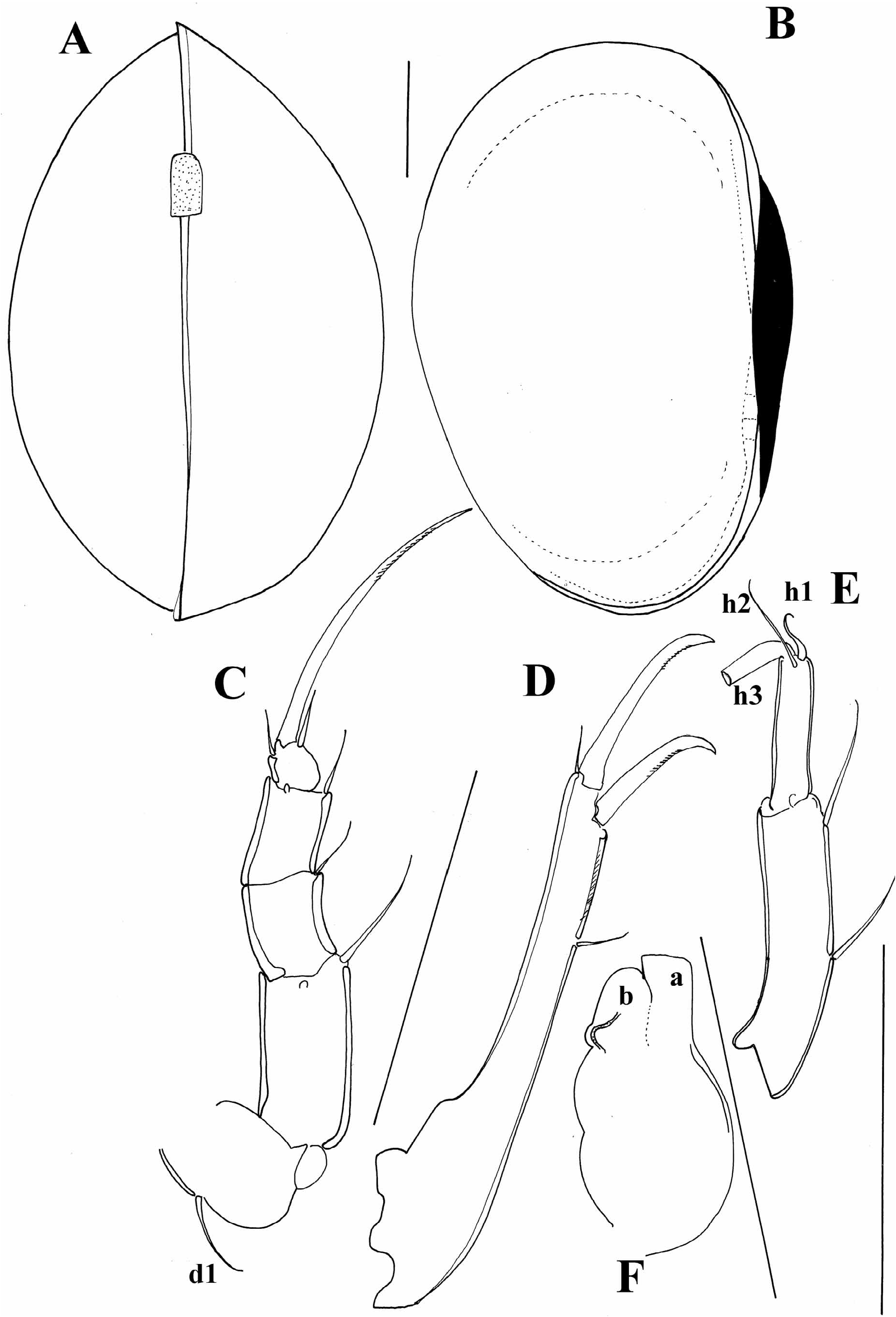
Two species have been described from South Africa: Cyclocypris pusilla Sars, 1895 and C. castanea Brady, 1913 , and they are the only two representatives of the genus found out of the Holarctic. Sars (1895) gave a short description and provided only two drawings of the carapace, according to which C. pusilla resembles a lot C. ovum ( Jurine, 1820) ( Figure 2A, B View FIGURE 2 ). Although the type material I have examined consists of a poorly preserved slide and two specimens in alcohol, there are a couple of differences that clearly separate C. pusilla from C. ovum . First of all, the terminal segment of the T3 is conspicuously elongated ( Figure 2E View FIGURE 2 ), but the h1 seta is, like in C. ovum , “S”- shaped. There are some differences in the appearance of the hemipenis, lobe “a” being much smaller in the South African species ( Figure 2F View FIGURE 2 ). The T2 ( Figure 2C View FIGURE 2 ) has the typical morphology of the subfamily, while the UR ( Figure 2D View FIGURE 2 ) is elongated and very similar to that of C. ovum . Cyclocypris castanea Brady, 1913 is not included in the key to species and is listed at the end of the paper. This species has conspicuously long marginal pore canals and a highly arched dorsal carapace margin, but further study of the material is important to properly evaluate this species. Klugh (1923) described two Cyclocypris species from Canada: C. castanea Klugh, 1923 and C. lutea Klugh, 1923 . The first one is a homonym, but since the taxonomic position of both Canadian species is doubtful, they are only listed at the end of the paper.
Cyclocypris helokrenica Fuhrmann & Pietrzeniuk, 1990 is very closely related to C. ovum , and although Meisch (2000) provides a couple of diagnostic characters for this species, his opinion is that “more research is needed to confirm or invalidate the taxonomic validity of these characters”. This species is also listed at the end of the paper and was not included in the keys. It needs to be mentioned here that species already synonymised in the more recent publications (i.e. Meisch, 2000) are not considered here, because I agree with the systematic decisions of their authors.
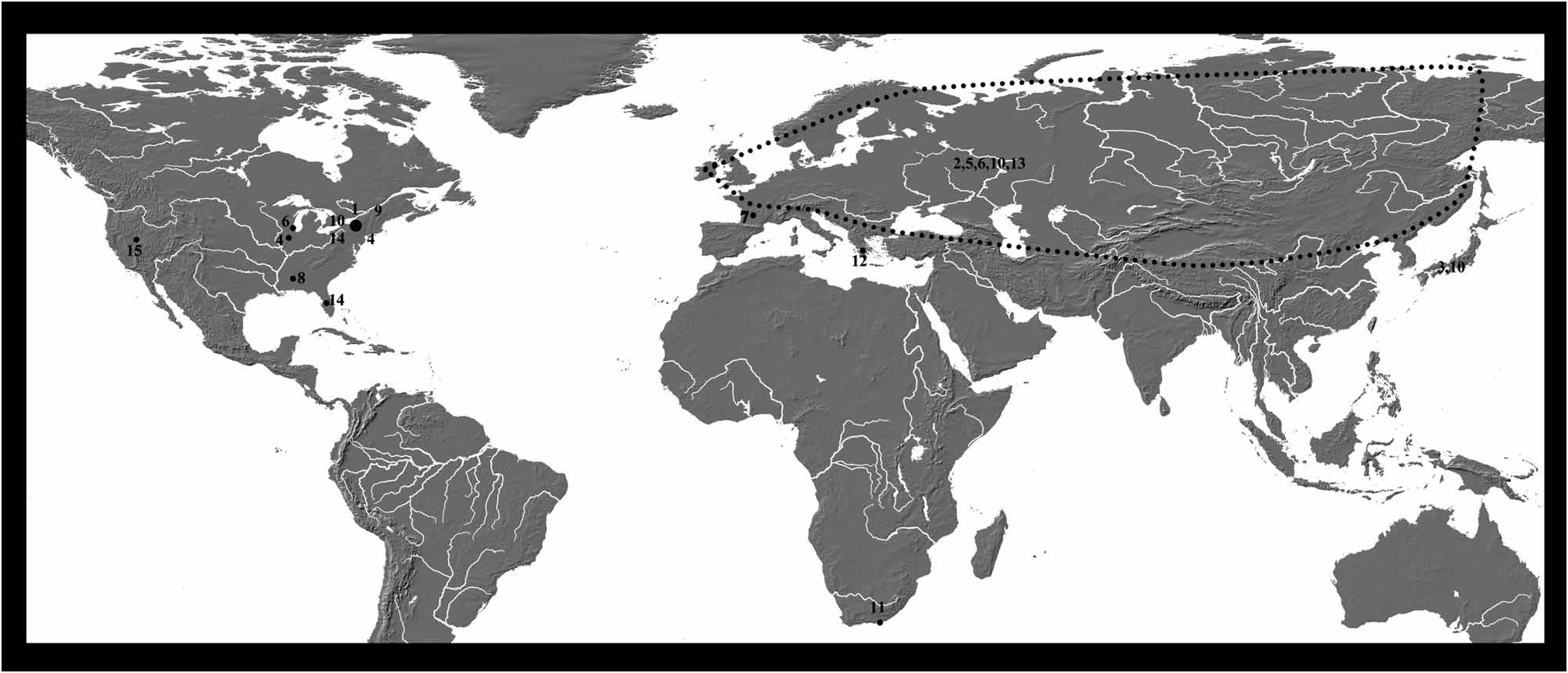
Distribution: Except for C. ovum and C. laevis , which are known from all around the Holarctic, other species have much more restricted distributions ( Figure 3 View FIGURE 3 ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Cyclocypris Brady & Norman, 1889
| Karanovic, Ivana 2011 |
Cypris (Leucocypris) drastichi Jančařík, 1949
| Jancarik 1949 |
Cyclocypria
| Dobbin 1941 |
Cyclocypria kincaidia
| Dobbin 1941 |