
Cymbasoma lourdesae, Suárez-Morales, Eduardo & Mckinnon, David, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4102.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9A7BA798-AA7C-4CAA-B42C-1E260CA573E4 |
|
DOI |
https://doi.org/10.5281/zenodo.6091301 |
|
persistent identifier |
https://treatment.plazi.org/id/03C4CA6D-D537-FF8D-FF12-546097C22D41 |
|
treatment provided by |
Plazi |
|
scientific name |
Cymbasoma lourdesae |
| status |
sp. nov. |
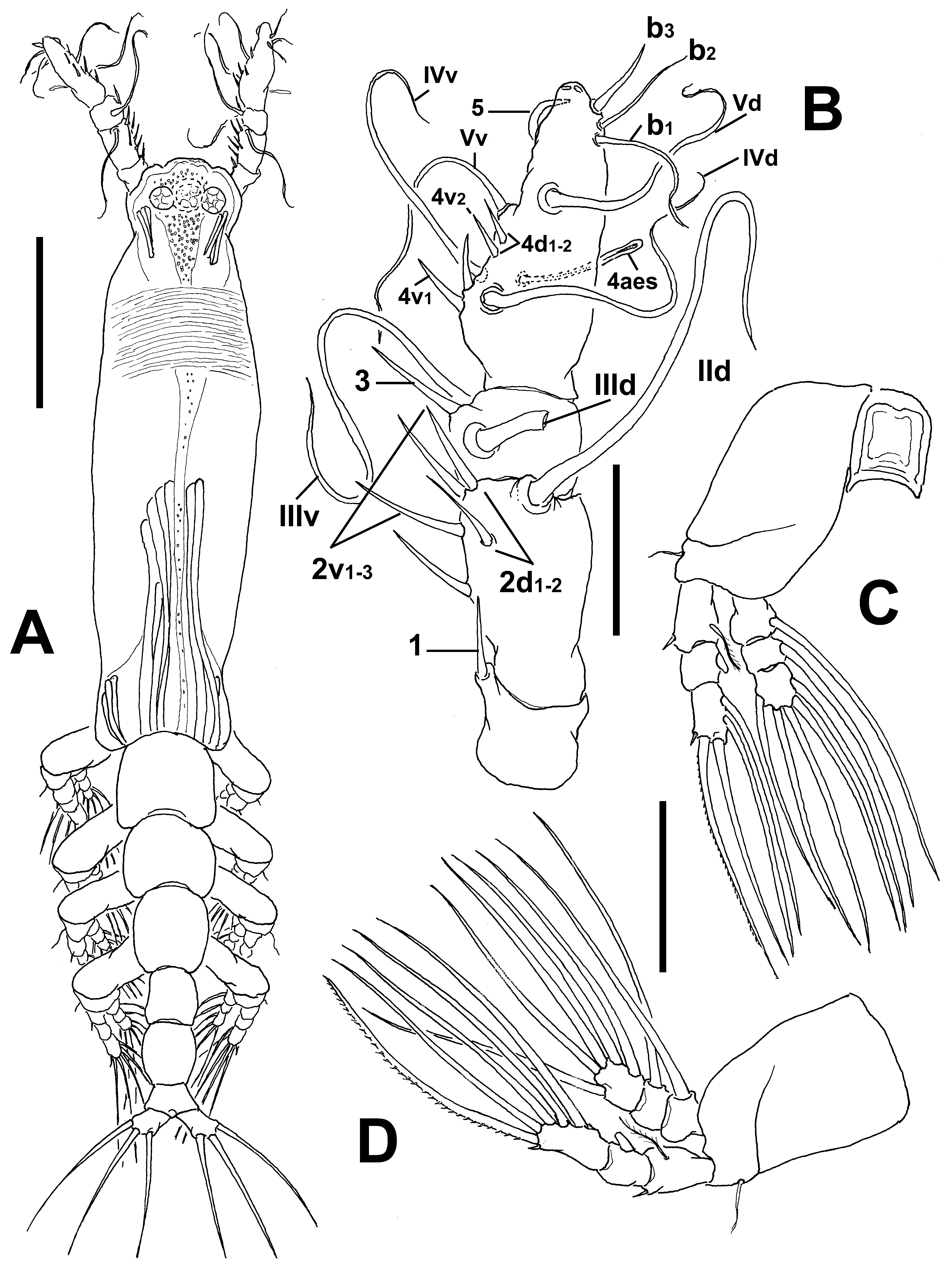
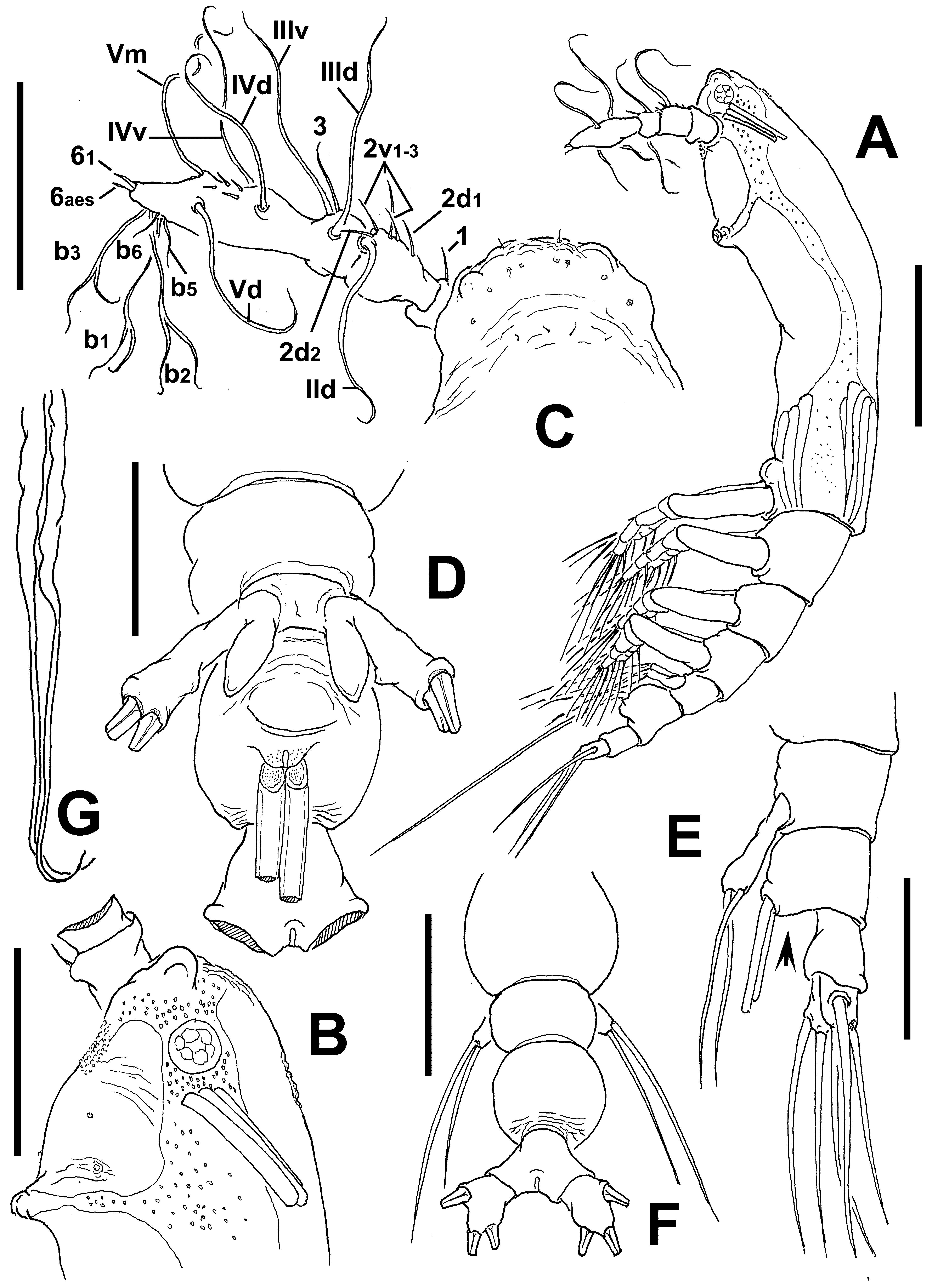
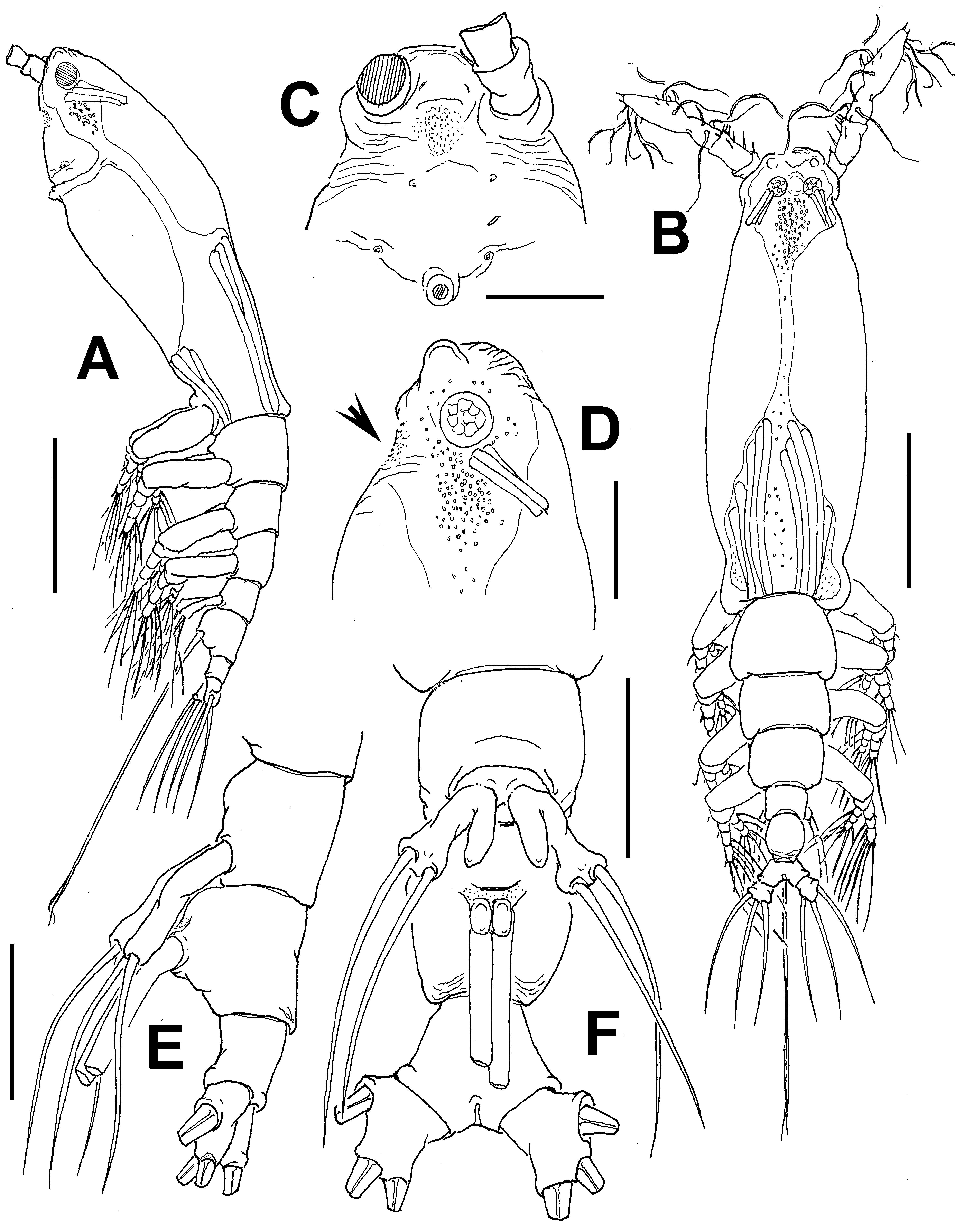
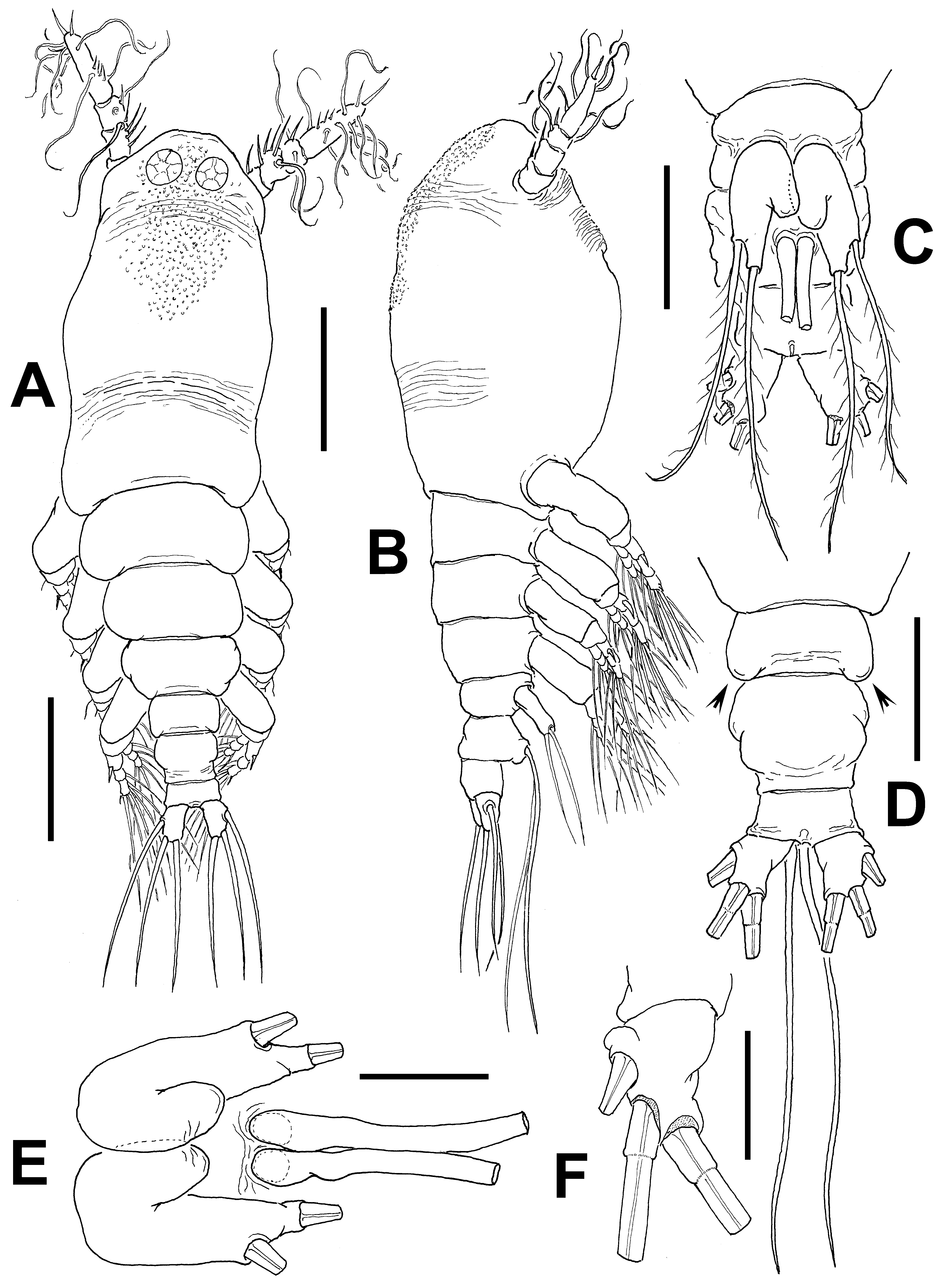
Cymbasoma lourdesae sp. nov.
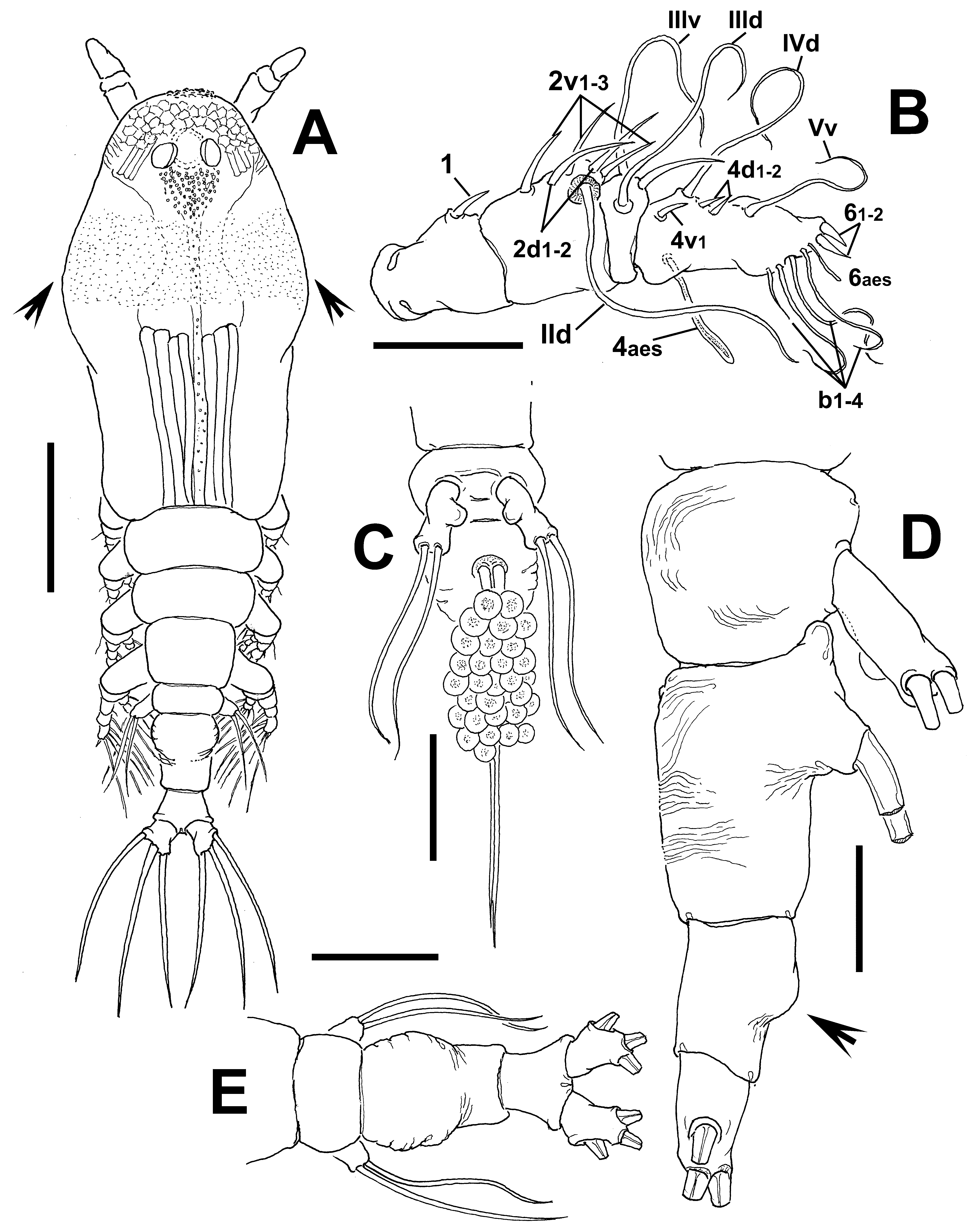
( Figs 24 View FIGURE 24 , 25 View FIGURE 25 )
Material examined. Adult female holotype and adult female paratype from Rhyll, Western Port Bay, Victoria, Australia (38°26.792’ S, 145°18.496’ E), partially dissected, parts mounted on slides in glycerine, sealed with Entellan®. Date of collection: 10th May 1983. Slides deposited in the collection of MTQ, Australia (cat. MTQ W34385, MTQ W34386).
Description of adult female. Body moderately robust ( Figs 24 View FIGURE 24 A, 25A, B); body length of holotype female 0.98 mm. Paratype female = 0.93 mm. Cephalothorax approximately 0.56 mm long, representing 59% of total body length. Midventral oral papilla protuberant, located at 31% of cephalothorax length. Pair of relatively small ocelli present, pigment cups weakly developed, separated by one eye diameter, lightly pigmented; ventral cup slightly larger than lateral cups. Frontal area ornamented with pattern of shallow transverse striations ( Fig. 25 View FIGURE 25 D); two pairs of frontal sensilla were observed ( Fig. 24 View FIGURE 24 C). Dorsal surface of cephalothorax smooth except for scattered transverse striae arranged in curved pattern; ventral surface ornamented with few transverse striae between antennule bases and oral area ( Fig. 25 View FIGURE 25 C). Additional ornamentation of ventral surface including: 1) medial field of cuticular papillae posterior to antennule bases, with striae at both sides ( Fig. 24 View FIGURE 24 B, arrowed in Fig. 25 View FIGURE 25 D); 2) pair of symmetrical, weakly developed nipple-like processes on anterior ventral surface located at both sides of oral papilla.
Urosome consisting of fifth pedigerous somite, genital double-somite and anal somite, together representing 14% of total body length. Relative lengths of urosomites (fifth pedigerous, genital double and free anal somites) 38.5: 41.4: 20.1 = 100, respectively ( Fig. 25 View FIGURE 25 E). Lateral margins of fifth pedigerous somite straight ( Fig. 25 View FIGURE 25 F). Genital double-somite longest of urosome, with smooth dorsal and ventral surfaces except for transverse wrinkles along posterior margins ( Figs 24 View FIGURE 24 D, 25F). Lateral margins of somite expanded into globose shape. Ventral surface of somite protuberant, with posterior margin produced ventrally, protruding from succeeding anal somite in lateral view (arrowed in Fig. 24 View FIGURE 24 E). Ovigerous spines paired, separated at base, relatively short, 43 % of total body length ( Fig. 24 View FIGURE 24 G). Spines slender, straight at their base and along shaft, with weak subterminal expansions and tapering apically; spines equally long, about 45% of total body length (0.4 mm) ( Figs 24 View FIGURE 24 A, 25A, B). Anal somite without medial constriction ( Figs 24 View FIGURE 24 D, F, 25F). Caudal rami divergent, subquadrate, about as long as wide, armed with three caudal setae ( Figs 24 View FIGURE 24 F, 25F).
Antennule length 0.22 mm, representing about 23.5% of total body length and 39% of cephalothorax length, 4- segmented, segments 3 and 4 partially fused. Relative length of distal antennulary segment 53%. In terms of pattern described by Grygier & Ohtsuka (1995) for female monstrilloid antennulary armature, short, spiniform element 1 present on first segment; elements on second segment: 2d1-2, 2v 1-3, and IId. Third segment with element 3 being slender, setiform, elements IIId and IIIv of normal aspect. Segment 4 bearing short, spiniform elements 4d1,2, 4v 1- 2, element 4v 3 not observed; setae IVd, IVv, Vd, Vm present. Element 5 absent. Subterminal elements b1-3, 5, 6 present, b1-3 dichotomously branched. Elements 61 and 6aes present ( Fig. 24 View FIGURE 24 C).
Incorporated first pedigerous somite and succeeding three free pedigerous somites each bearing a pair of biramous legs. Pedigerous somites 2–4, together accounting for 26% of total body length. Legs 1–4 as in C. pseudoquadridens except for smooth intercoxal sclerites.
Fifth legs medially conjoined, bilobate, outer (exopodal) lobe cylindrical, distally truncate. Lobe with low inner protuberances on middle inner margin, armed two equally long apical setae ( Figs 24 View FIGURE 24 D, 25F). Inner lobe thumb-like, shorter than outer lobe, reaching about half the length of exopodal lobe, unarmed.
Male: unknown.
Type locality. Rhyll, Western Port Bay, Victoria, Australia (38°26.792’ S, 145°18.496’ E).
Etymology. The species is dedicated to Lourdes Vásquez-Yeomans, a researcher at El Colegio de la Frontera Sur, for her long work in the study of the tropical marine zooplankton and ichthyoplankton.
Diagnosis. Cymbasoma with cephalothorax, representing 59% of total body length, third and fourth antennulary segments fused. Ventral field of papilla-like processes on cephalic area between antennule bases and oral papilla. Genital double-somite with moderately expanded margins, with wrinkles on posterior ventral margin. Anal somite not constricted. Fifth leg with elongate, cylindrical outer lobe with two distal setae; inner lobe welldefined, arising basally and reaching about half the length of outer lobe, unarmed.
Remarks. This is another species with only two distal setae on the outer lobe of the fifth leg. It differs from its Australian congeners sharing this peculiar character ( C. dakini sp. nov., C. agoense , C. tharawalorum sp. nov.) in the length of the inner lobe, which is shorter than in C. dakini , not reaching the distal margin of the outer lobe. The shape of the genital double-somite is also different in these species; the new species has a globose shape ( Figs 24 View FIGURE 24 D, F, 25F) thus diverging from the moderately produced genital double-somite in C. dakini . Also, some details of the antennulary armature are useful to distinguish these species; in C. dakini element 5 on the fourth segment is strong, spiniform, curved ( Fig. 6 View FIGURE 6 B) whereas it is absent in the new species. On the same segment, the elements of group “b”, b1-3 are short, unbranched in C. dakini ( Fig. 6 View FIGURE 6 B) but they are long and dichotomously branched in the new species. Most importantly, the antennule of the new species has segments 3–4 fused, thus diverging from the clearly separated antennulary segments 3–4 in C. dakini . Also, the new species has a field of papilla-like elements on the ventral surface of the cephalic area ( Fig. 25 View FIGURE 25 C, D), a character absent in the other species. The new species differs from both C. tharawalorum in the body shape; the latter species is clearly more robust. The shape of the double genital somite differs in these two species; it is strongly globose in the new species and in C. tharawalorum only the proximal half is expanded and bears lateral striae, and the posterior half is straight ( Fig. 65 View FIGURE 65 D, E). The shape and relative size of the antennules are also different in these species; it is short, with the last segment representing less than 40% of the antennule length vs. 53% in the new species. In C. tharawalorum the fifth leg inner lobe is small, globose and inserted medially ( Fig. 65 View FIGURE 65 C) whereas it arises proximally and it is oblong in the new species. It differs in several characters from C. agoense , that also has two setae on the outer lobe; in C. agoense the outer lobe is strong, wide ( Sekiguchi 1982: fig. 6F; Fig. 56 View FIGURE 56 E), thus contrasting with the elongate outer lobe of the new species; a similar difference is observed for the inner lobe. The body of C. agoense is short, robust ( Sekiguchi 1982: fig. 6A; Fig. 56 View FIGURE 56 A) and clearly differs from that of the new species. In addition, the globose genital double-somite of the new species diverges from the quadrate somite present in C. agoense ( Sekiguchi 1982: fig. 6A; Fig. 56 View FIGURE 56 D). The antennule armature includes some additional differences; setae of the “b” group are long and distally branched in the new species vs. relatively short, unbranched “b” setae in C. agoense ; also, the two apical spiniform elements 61-2 are remarkably long in C. agoense ( Sekiguchi 1982: fig. 6C) whereas they are clearly shorter in the new species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |