
Cyclops lacustris G.O. Sars, 1863
|
publication ID |
https://doi.org/ 10.5852/ejt.2016.212 |
|
DOI |
https://doi.org/10.5281/zenodo.5619469 |
|
persistent identifier |
https://treatment.plazi.org/id/A0385210-FF8F-FFB1-256F-FAA0FBD8766A |
|
treatment provided by |
Plazi |
|
scientific name |
Cyclops lacustris G.O. Sars, 1863 |
| status |
|
Cyclops lacustris G.O. Sars, 1863
Figs 7 View Fig ̅10
Cyclops lacustris G.O. Sars, 1863: 30
Cyclops lacustris – Sars, 1914: 35 ̅36, pl. 18. — Rylov 1948: 187 ̅189, fig. 38, tab. 13. — Lindberg 1957: 85 ̅87, figs 90̅94, tab. 13. — Einsle 1993: 141 ̅142, fig. 84; 1996a: 41̅43, fig. 26. — Deimantovica 2010: 216 ̅222, figs 2̅5, tabs 1, 3–4.
Cyclops strenuus f. lacustris – Lilljeborg 1901: 31 ̅32, pl. 2, fig. 23.
Type material
Lectotype (designated here)
SOUTHERN NORWAY: ♀, dissected on two slides labelled as “ lectotypus b” and “ lectotypus c”, Lake Mjøsa , 60º42' N, 11º1' E, 121 m a.s.l., surface area 369.2 km 2, max. depth 443 m, G.O. Sars coll., prep. Fr. Kiefer, 27 Aug. 1971 ( ZMO).
GoogleMapsParalectotypes
SOUTHERN NORWAY: 1 undissected ♀ labelled as “ lectotypus a”, 1 Ƌ labelled as “ lectotypus d”, same locality as in lectotype, G. O. Sars coll., prep. Fr. Kiefer, 27 Aug. 1971 (both ZMO); 2 ♀♀ and 1 Ƌ dissected by the authors from vial “Sars Coll. 12977” ( ZMO), and 1 ♀ and 1 Ƌ dissected by the authors from vial “Sars Coll. 12976” ( ZMO); two antenna of single ♀, “ Cyclops lacustris ♀ 2 A 2 Kiefer 26.4.83, Norwegen, Mjösen 4894, Oslo Mus. F 12977” ( SMNK:11366); 1 undissected ♀ “ Cyclops lacustris Sars , ♀ Kiefer 31 Aug. 71, Norwegen Mjøsen” ( SMNK: 9567); and 2 ♀♀ dissected on two slides “ Cyclops lacustris Sars , ♀, C. Kiefer 31 Aug. 71, Norwegen Mjøsen” ( SMNK: 9568 (Abd+ P 5), SMNK: 9569 ( A 1- P 4).
Other material examined
NORWAY (southern): 1 ♀, Øymarksjøen, 59º23' N, 11º39' E, 107 m a.s.l., surface area 14.3 km 2, max. depth 37.6 m, 18 Jul. 2012, leg. I. Dimante-Deimantovica & B. Walseng ( MIZ 2/2016/16); 1 ♀, Store Le, 59º17' N, 11º48' E, 102 m a.s.l., located on the border between Norway and Sweden, surface area 136.1 km 2, of which 14.88 km 2 in Norway, max. depth 58.2 m, 2011, P.2872, leg. I. Dimante-Deimantovica & B. Walseng ( MIZ 2/2016/17); 1 ♀, Aremark, 59º15' N, 11º40' E, 105 m a.s.l., surface area 7.47 km 2, max. depth 39.5 m, 18 Jul. 2012, leg. I. Dimante-Deimantovica & B. Walseng ( MIZ 2/2016/18); 2 ♀♀, 1 Ƌ, Aspern, 59º9' N, 11º43' E, 105 m a.s.l., surface area 6.76 km 2, max. depth 46.5 m, Jul. 2011, leg. I. Dimante-Deimantovica & B. Walseng ( MIZ 2/2016/19-20 and MIZ 2/2016/21, respectively).
SWEDEN: 4 ♀♀, K. Lindberg collection Lund, jar nr. 109, S. 28 ( ZMLU).
LATVIA: 1 ♀ dissected ( MIZ 2/2016/22) and 1 ♀ in alcohol ( IDD), 1 CV Ƌ parts ( A 2, Mxl, P 1) ( MIZ 2/2016/23), 2 CIV ♀♀ parts ( A 1, A 2, Mxl, P 1) ( MIZ 2/2016/24-25), 4 CIV parts ( A 2, Mxl, P 1) ( MIZ 2/2016/26-29), Lake Svente, 55º51' N, 26º22' E, southeastern part of Latvia, 137 m a.s.l., surface area 7.35 km 2, max. depth 38 m, 30 Aug. 2007, leg. I. Deimantovica.
Description
Female
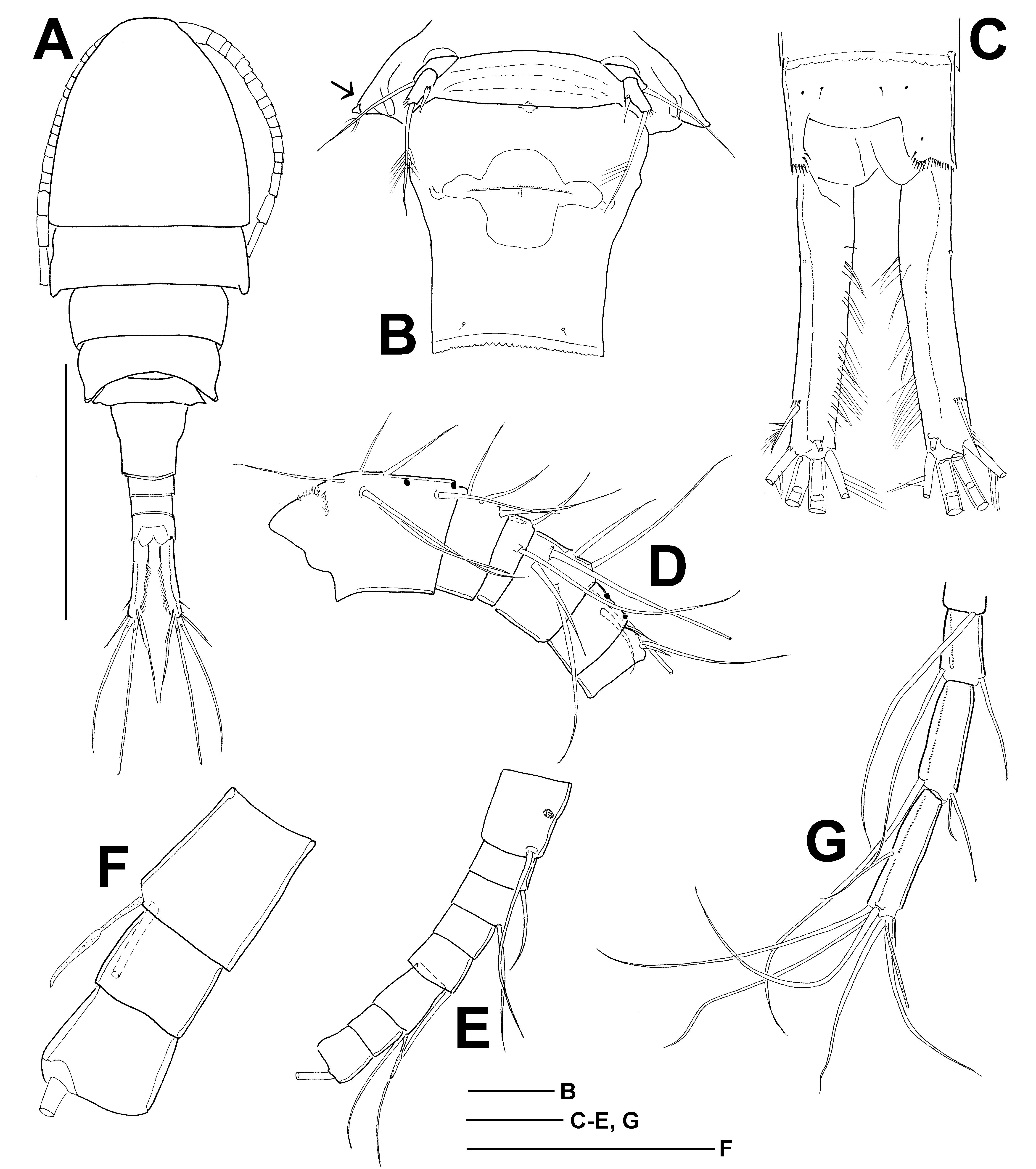
Body length 1375–1800 µm (the female here designated as lectotype was originally dissected by F. Kiefer, and no data are available on the morphometric traits of the prosome and the total body length). Cephalothorax length/width 1.1̅1.3, cephalothorax width/genital double-somite width 2.6̅2.8,
prosome length/urosome length 1.4̅1.7. Pediger 2 ( Fig. 7A View Fig ) without distinct posterolateral lobes, and not wider than succeeding somites. Pediger 5 not protruding laterally ( Fig. 7B View Fig vs. Fig. 1B View Fig ).
Genital double-somite ( Fig. 7B View Fig ) longer than its greatest width. Seminal receptacle ( Fig. 7B View Fig ) as typical in genus, transverse ridge present next to copulatory pore. Posterior margin of anal somite with continuous row of spinules, anal sinus without surface ornamentation, anal operculum weakly developed. Caudal rami ( Fig. 7C View Fig ) length/width 4.3̅7.5 (lectotype, 4.9), medial margin bearing hairs. Midline crest running almost full length of ramus on dorsal surface. Few spinules present on lateral margin in anterior third, indicating presumptive insertion site of anterolateral accessory seta (I). Spinules also present at insertion of caudal setae II and III. Caudal seta II inserted at distance of 0.20̅0.26 ramus length measured from posterior end (lectotype, 0.25). Relative length of caudal setae VII, VI, V, IV, and III: 1.2̅2.0 (lectotype, 1.5), 1.8̅2.6 (lectotype, 2.3), 3.7̅6.0 (lectotype, 5.3), 3.2̅4.9 (lectotype, 4.0), 1.0. Seta VI (medialmost) 1.0̅1.4 (lectotype, 1.35) times as long as caudal rami, seta V (longest) 0.63̅1.0 (lectotype, 0.95) times as long as urosome. Caudal setae homonomously setulose, setae IV and V with breaking plane.
Antennule reaching middle of pediger 3, segmentation and setation pattern as in C. bohater . Aesthetasc on segment 12 ( Fig. 7D View Fig ) reaching middle to distal margin of segment 14; aesthetasc length 2.9̅3.6% of body length (n = 10; no data on lectotype). Aesthetasc on segment 16 reaching near insertion site of medial seta of segment 17 (approx. middle of terminal segment); aesthetasc length 2.6̅3.6% of body length (n = 9; no data on lectotype). Spinules only present on ventral surface of segment 1. Last three segments of antennule with hyaline membrane.
Antenna composed of coxobasis and three-segmented endopodite, and bearing 3, 1, 9 and 7 setae, respectively. Inner medial seta of coxobasis ( Fig. 7E View Fig ̅F) with long setules, outer medial seta naked. Exopodite seta reaching beyond enp3 and bearing setules conspicuously longer in proximal than in distal section of seta. On frontal surface of coxobasis, spinules (4, 5) present in ~4/10 of segment and group of longer spinules appear more proximally next to lateral margin, few long spinules usually present in oblique/longitudinal row near middle line in proximal third of segment (for similar pattern see Fig. 2A View Fig ). Caudal surface of coxobasis ( Fig. 7E View Fig ̅F) with double row of long spinules near long proximal spinules on lateral margin, long and thin spinules (5̅7) in longitudinal row near lateral margin, and oblique field of small spinules proximally to insertions of medial setae.
Labrum with distal teeth in arc, lateral lobe with small spinules. Distal hairs long and arranged in two groups. Epistoma and vertical cleft naked. Segmentation and setation of mandible, maxillule, maxilla and maxilliped as in C. bohater . No cuticular ornamentation present next and proximally to mandibular palp. Maxillulary palp usually without spinules ( Fig. 7G View Fig ) (spinules absent in lectotype). Proximal seta and at least one (sometimes two or all three) of lateral lobe setae of maxillulary palp with distinctly long setules ( Fig. 7G View Fig ̅H). Claw-like attenuation of basipodite of maxilla with spinules on both concave (inner) and convex (outer) margin. Basipodite seta inserted in front of claw-like attenuation with long setules on both concave and convex edge in proximal half, and tiny spinules on convex edge in distal half (for similar pattern see Fig. 3A View Fig ). Maxilliped: first endopodal segment bearing long and thin spinules on frontal surface; basipodite with spinules arranged in two groups on caudal surface and long and thin spinules on frontal surface; syncoxopodite frontally adorned with small spinules midway between median and distalmost setae, tiny spinules sometimes present ( Sweden, ♀ -2) near finger-like membraneous element (for similar pattern see Fig. 3B View Fig ).
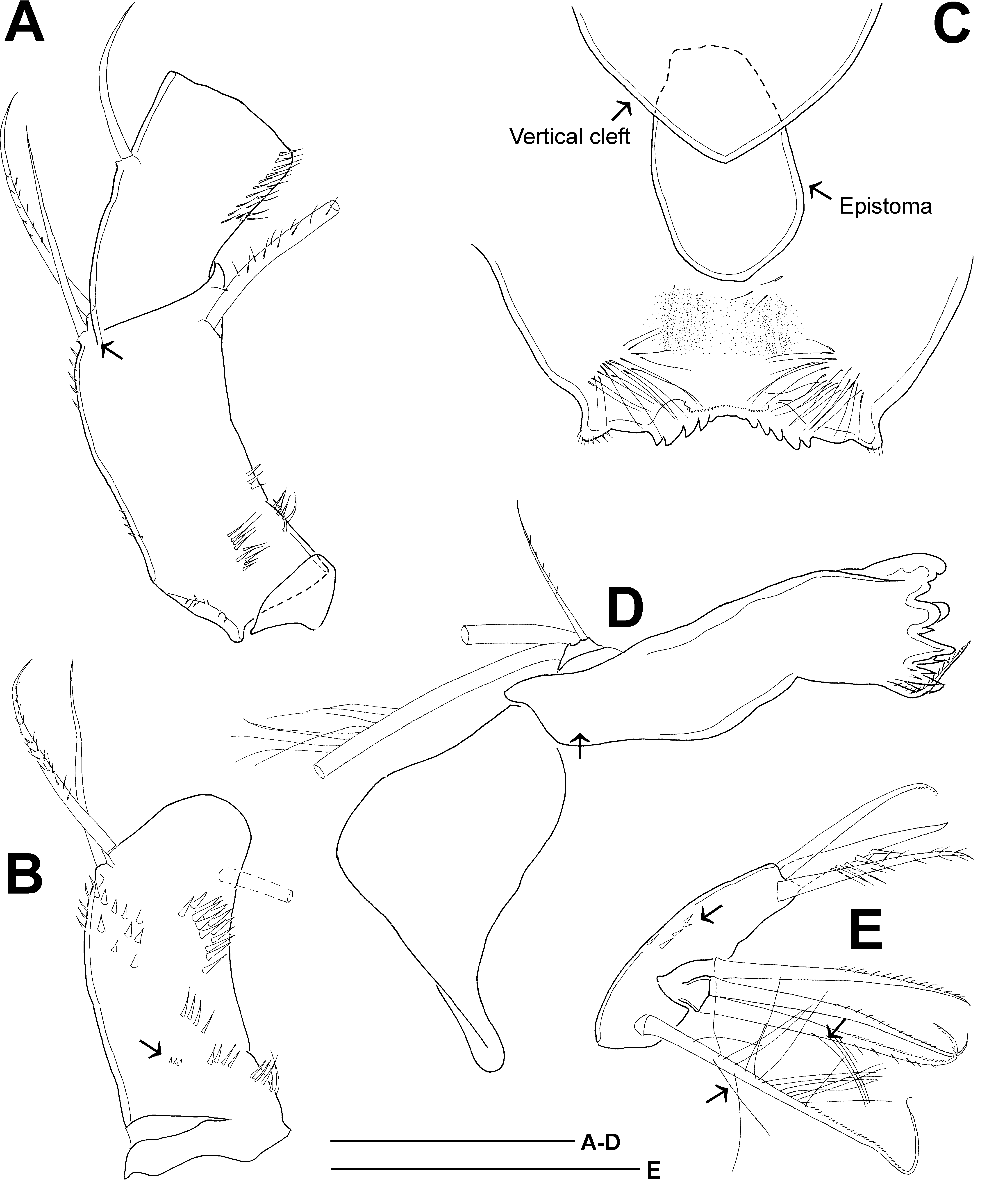
P1̅P4 setation formula same as in C. bohater ( Table 2). Medial spine of P1 basipodite ( Fig. 8A View Fig ): setules distinctly longer near base of seta and becoming abruptly shorter towards distal end, or length of setules almost gradually decreasing (lectotype). Medial spine reaching beyond distal margin of enp2 up to middle of enp3. Long spinules arranged in arc on frontal surface of P1 basipodite between insertions of exo- and endopodite. Intercoxal sclerites naked on frontal and caudal surfaces in P1̅P3, and sparsely pilose on caudal surface in P4. Obtuse protuberances of P4 intercoxal sclerite low, slightly extending beyond distal margin of segment ( Fig. 8B View Fig ). Medial expansion of basipodite usually pilose in P1̅P3 and naked in P4; basipodite sometimes naked in P2̅P4 (Sars Coll. 12977 ♀ -1) or pilose in all swimming legs ( Sweden, ♀ -2). Caudal surface ornamentation of P4 coxopodite ( Fig. 8B View Fig ) composed of several groups of spinules (groups are coded according to Einsle 1996a): A – with 6–9 spinules in medial section and 5–10 spinules in lateral section; B – few spinules (5), group sometimes absent; C – 5–7 spinules, sometimes accompanied with 2–4 spinules directly above former row; D – usually single spinule, sometimes group of spinules (7 + 1); E – 5–8 spinules. P4 coxopodal seta conspicuously longer (1.6̅2.1×; lectotype, 1.6×) than height of medial expansion of basipodite. On caudal surface, distal margin of first and second exopodal segment of P4 naked or bearing spinules. P4 enp3 2.8̅3.8 (lectotype, 3.0) times as long as wide; of apical spines medial one 2.2̅3.8 (lectotype, 3.4) times as long as lateral and 0.8̅1.1 (lectotype, 0.95) times as long as segment.
P5 ( Fig. 7B View Fig ) segmentation and setation as typical in genus. Distal segment 1.9̅2.3 times as long as wide. Medial spine inserted near half-length of segment (0.47̅0.64 segment length measured from proximal margin; 0.60 segment length in lectotype), medial spine 0.33̅0.62 (lectotype, 0.37) times as long as distal segment. Lateral seta on proximal segment relatively long, 0.70̅0.90 (lectotype, 0.70) times as long as apical seta and 2.5̅4.0 (lectotype, 2.5) times as long as distal segment. Apical seta 3.3̅4.6 (lectotype, 3.6) times as long as distal segment. Spinules present at insertion of median spine and apical seta, but absent at insertion of lateral seta.
Male
Body length 1135–1325 µm, cephalothorax length/width 1.3–1.4, prosome length/urosome length ca 1.6. Pediger 5 laterally not protruding. Caudal rami ( Fig. 8C View Fig ) without dorsal crest, 3.5̅5.1 times as long as wide, pilose on medial margin. Short transverse row of spinules present in anterior third at presumptive insertion site of ancestral anterolateral accessory seta (I). Spinules also present at insertion of caudal seta II and III. Caudal seta II inserted at distance of 0.26̅0.30 ramus length measured from posterior end. Relative length of caudal setae VII, VI, V, IV, and III: 1.8̅2.5, 2.3̅2.8, 4.1̅5.4, 3.3̅4.4, 1.0. Seta VI 1.5̅1.7 times as long as caudal rami, seta V 0.81̅0.98 times as long as urosome.
Antennule 17-segmented, but two terminal segments (ancestral segment XXV and compound segment XXVI̅XXVIII) fused on ventral surface. Armature formula as in C. bohater . Antenna segmentation and setation as in female, except for enp2, which has 8 setae. Spinule ornamentation of antennal coxobasis ( Fig. 8E View Fig ) similar to that in female, inner medial seta bearing long setules, outer seta naked. Mouthparts similar to those in female. Lateral lobe setae of maxillulary palp sometimes lacking long setules ( Fig. 8F View Fig ). Maxilliped syncoxopodite bearing small spinules midway between median and distalmost setae (verified in male from Lake Aspern, Norway).
P1̅P4 segmentation and setation as in C. bohater ( Table 2.). Medial spine of P1 basipodite usually with homonomous setulation ( Fig. 9A View Fig ) and exceeding distal margin of enp2; distinctly long setules sometimes present near base of seta (Lake Aspern, Norway). Long spinules arranged in arc on frontal surface of P1 basipodite between insertions of exo- and endopodite. Lateral spine of P2 exp1 more slender than other spines of exopodite, bearing long setules. Intercoxal sclerites naked in P1̅P3 and sparsely pilose on caudal surface in P4. Caudal surface ornamentation of P4 coxopodite composed of groups A, C and E; group B sometimes present (Lake Aspern, Norway) ( Fig. 9B View Fig cf. Fig. 8B View Fig ). Coxopodal seta ca 1.5× longer than height of medial expansion of basipodite. Medial expansion of basipodite apically pilose in P1̅P3, naked (paralectotype ̅ Sars Coll. 12976) or pilose (paralectotype ̅Sars Coll. 12977; Lake Aspern, Norway) in P4. P4 enp3 2.7̅3.3 times as long as wide. Of apical spines medial one 2.5̅3.7 times as long as lateral spine, as long or slightly (~1.1×) longer than segment.
P5 ( Fig. 8D View Fig ) distal segment 1.6̅1.9 times as long as wide, medial spine inserted near half-length of segment (0.5̅0.6 segment length measured from proximal margin), medial spine 0.41̅0.50 times as long as distal segment. Lateral seta on P5 proximal segment 0.60̅0.74 times as long as apical seta and 3.0̅4.2 times as long as distal segment. Apical seta 5.0̅6.0 times as long as distal segment. Spinules present at insertion of median spine and apical seta, but absent at insertion of lateral seta.
P6 ( Fig. 8D View Fig ) composed of three elements; median and lateral seta 1.1̅1.4 times and 3.0̅4.3 times as long as median spine, respectively. P6 flap naked.
Morphology of the species diagnostic characters in the late copepodid instars (Lake Svente, Latvia)
Antennule segmentation and setation in CIV instar as in C. bohater . Inner medial seta of antennal coxobasis with conspicuously long setules, outer medial seta naked in CIV and CV; second endopodal segment with 7 and 8 setae in CIV and CV (male), respectively. Long setules present on proximal seta of maxillulary palp and at least one seta of lateral lobe of maxillulary palp both in CIV female and CV male. Setulation of medial spine of P1 basipodite homonomous or heteronomous (few longer setules present proximally) in CIV, and heteronomous in CV male (verified in single specimen).
Remarks
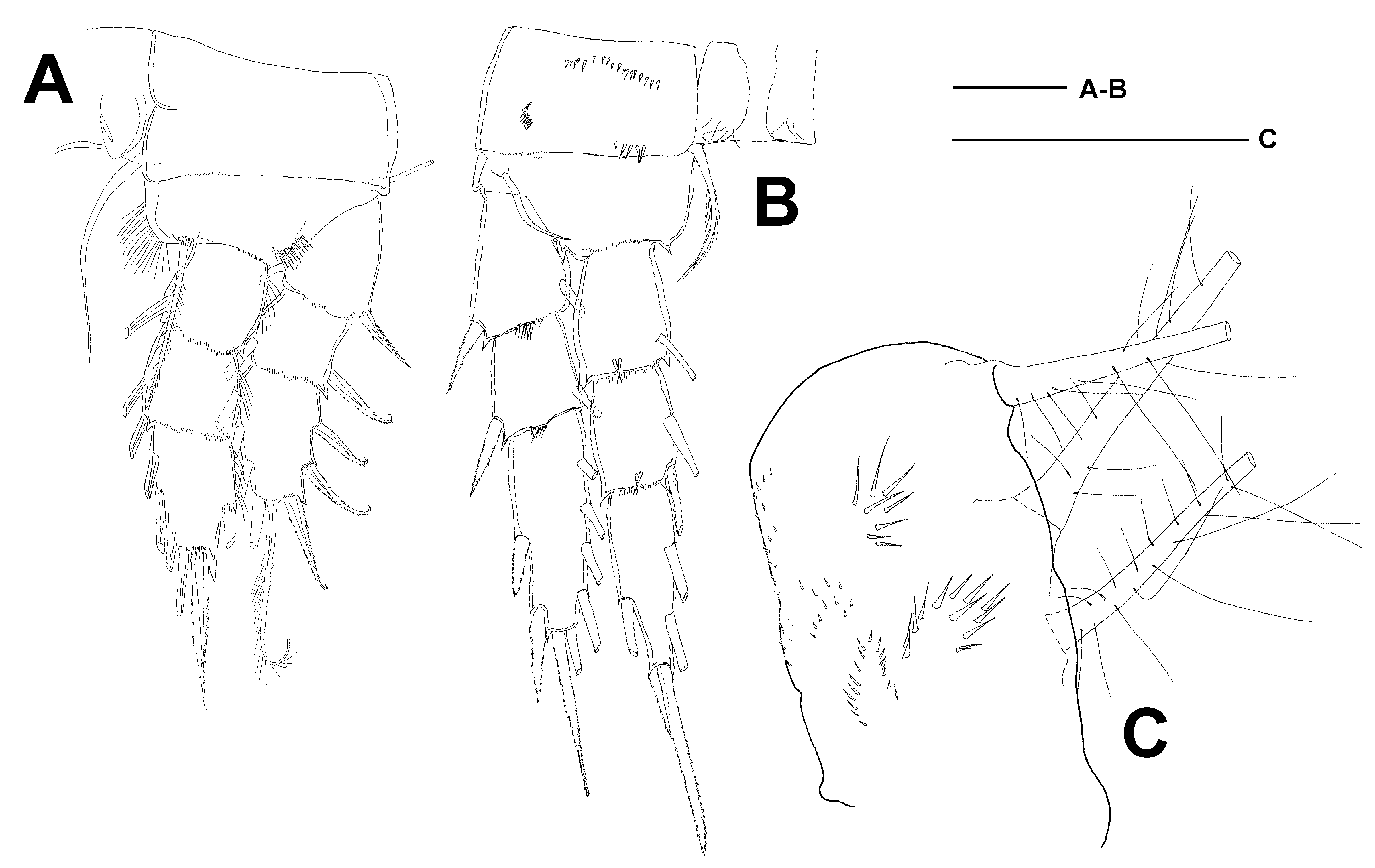
The species was originally described from two large lakes (Mjøsa (or Mjøsen) and Tyrifjord) in Norway; a holotype was not designated by the author ( Sars 1863). The Zoological Museum in Oslo holds the syntypes of C. lacustris collected by Sars from Lake Mjøsa, comprising alcohol material and few microscope slide preparata made by F. Kiefer; the fate of the original material from Tyrifjord is unknown to us. Other parts of the Sars legacy, such as notes, manuscript drafts and unpublished drawings, are kept in the archives of the National Library of Norway. The first description of the species was part of a two-volumed hand-written manuscript (archive item Ms. Fol. 1109: 613) entitled “Om de i Christiania’s Omegn forekommende Ferskvandskrebsdyr. I og II” (On the freshwater crustaceans occurring in the vicinity of Christiania) and devoted to the Cladocera, Copepoda and Ostracoda; this work as a whole has never been published. In the archives we did not manage to identify the drawings of C. lacustris which Sars might have added to the above mentioned manuscript; in any case, the published description of the species ( Sars 1863) did not contain illustrations. Nevertheless, the published diagnosis, the illustrations that later appeared in the monograph of the Crustacea of Norway ( Sars 1914) and some fine pencil drawings of C. lacustris from Lake Mjøsa ( Fig. 10A View Fig ̅C; date of origin of these drawings is unknown), deposited in the archives, allow us to identify the species with great certainty.
Kiefer (1978) provided drawings of the female habitus and the third endopodal segment of leg 4 of the “ lectotypus ” of C. lacustris , which most likely show one of the two females that he selected from among the syntypes as “ lectotype ” in the early seventies. These two females (one dissected on two slides, and one undissected specimen) plus one male, all of which were labelled by Kiefer as “ lectotypus ”, are deposited in the Zoological Museum in Oslo. As the lectotype designation by Kiefer is invalid (only one specimen can be choosen as the lectotype, ICZN art. 74.1), we designated the lectotype of C. lacustris herein, which is the dissected female labelled by Kiefer as lectotypus “B” (containing Abd+P5) and “C” (containing A1̅P4).
Ecology
Cyclops lacustris is a true limnetic species, mostly known from comparatively large and deep oligotrophic to slightly eutrophic lakes. Adult individuals are generally found in the epilimnion, close to the surface ( Sars 1914); however, the species has also been recorded down to 20 m depth and deeper ( Rylov 1948). In Lake Mjøsa a significant part of the population has been found in deep waters. The vertical distribution depends on the season and developmental stage ( Huitfeldt-Kaas 1946; Jarl Eivind Løvik, unpublished data).
In Norway C. lacustris often co-occurs with typical glacial relict species ( Spikkeland et al. 2012). In 2006, when C. lacustris was evaluated for the Norwegian Red List database, it was only known from Lake Mjøsa and got included in the Red List as a critically endangered species, threatened by eutrofication (incl. oxygen decrease) and water temperature increase due to the global warming. Later, by applying new micro-characters in species identification, C. lacustris was discovered in several other lakes in SE Norway. The new finds indicated that the species had a wider distribution in the country than was thought before. At the moment the Red List status of C. lacustris in Norway has been changed from endangered to least concern (Bjørn Walseng, personal communication). The example of C. lacustris illustrates how alpha taxonomy may contribute to a better understanding of the actual geographic distribution of and environmental threats to a species.
Distribution
Verified occurrences from Northern Europe ( Norway, Sweden, Latvia) ( Table 5). For an overview of the literature data on the geographic distribution of C. lacustris , see the Discussion.




| ZMO |
Zoology Museum, Oxford University |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| O |
Botanical Museum - University of Oslo |
| A |
Harvard University - Arnold Arboretum |
| F |
Field Museum of Natural History, Botany Department |
| SMNK |
Staatliches Museum fuer Naturkunde Karlsruhe (State Museum of Natural History) |
| C |
University of Copenhagen |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| N |
Nanjing University |
| E |
Royal Botanic Garden Edinburgh |
| I |
"Alexandru Ioan Cuza" University |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| S |
Department of Botany, Swedish Museum of Natural History |
| ZMLU |
Lunds Universitet, Zoologiska Institutionen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
SubPhylum |
Crustacea |
|
Class |
|
|
SubClass |
Copepoda |
|
Order |
|
|
Family |
|
|
Genus |
Cyclops lacustris G.O. Sars, 1863
| Hołyńska, Maria & Dimante-Deimantovica, Inta 2016 |
Cyclops lacustris
| Deimantovica I. 2010: 216 |
| Einsle U. 1993: 141 |
| Lindberg K. 1957: 85 |
| Rylov V. M. 1948: 187 |
| Sars G. O. 1914: 35 |
Cyclops strenuus
| Lilljeborg W. 1901: 31 |
Cyclops lacustris G.O. Sars, 1863: 30
| Sars G. O. 1863: 30 |