
Cryptodesmus ellipsoideus Novis & Visnovsky, 2012
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.39.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.4926571 |
|
persistent identifier |
https://treatment.plazi.org/id/0391A571-2107-C660-78AC-FC984EA4FF50 |
|
treatment provided by |
Felipe |
|
scientific name |
Cryptodesmus ellipsoideus Novis & Visnovsky |
| status |
gen. et sp. nov. |
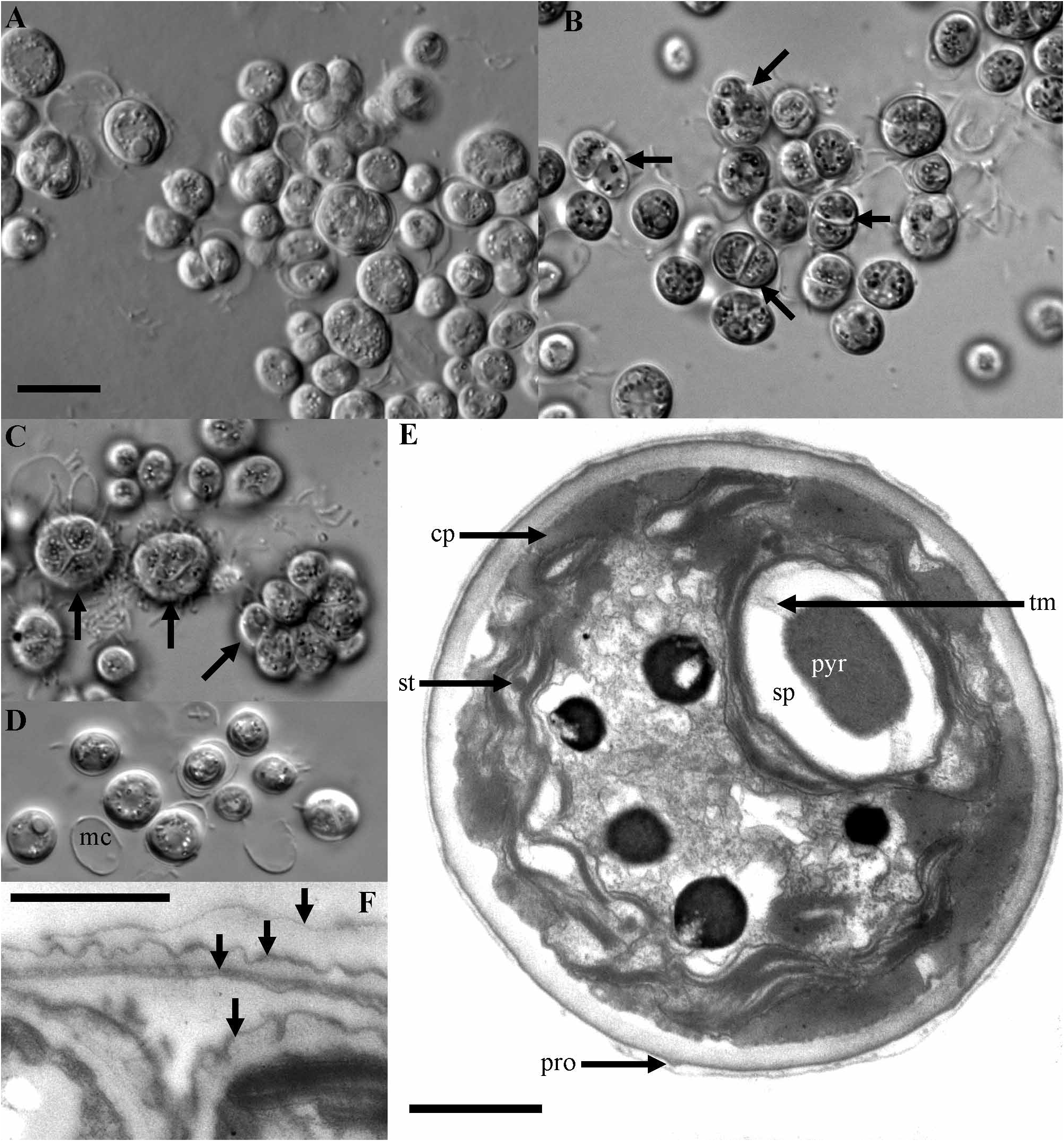
Cryptodesmus ellipsoideus Novis & Visnovsky , gen. et sp. nov. ( Figs 3A–F View FIGURE 3 )
Cellulae ellipsoideae vel subirregulares, 6.2–9.2 µm longae, 4.6–8.5 µm latae, unicae vel in greges aggregatae. Paries cellularis in LM patefactus laevis, tenuisque. Chloroplastus cupulatus, parietalis, pyrenoide unica prominenti per membranas thylakoidium unicas vel plures transversa atque in vagina amylacea segmentata e partibus duabus vel pluribus composita includenti. Parietes cellulares in TEM praecipue in cellulis dividentibus plicis irregularibus convolutis instructi, imprimis in stratis exterioribus. Autosporis 2–4–8(–16) per sporangium, saepe arcte contiguis, regenerans; cellulae reliquae matricales saepe intra cellulas cultas visae.
Type:— NEW ZEALAND: Westland : Mt Philistine, 1400 m, preserved cultured specimen from sample collected 30 November 2007, CHR610486 .
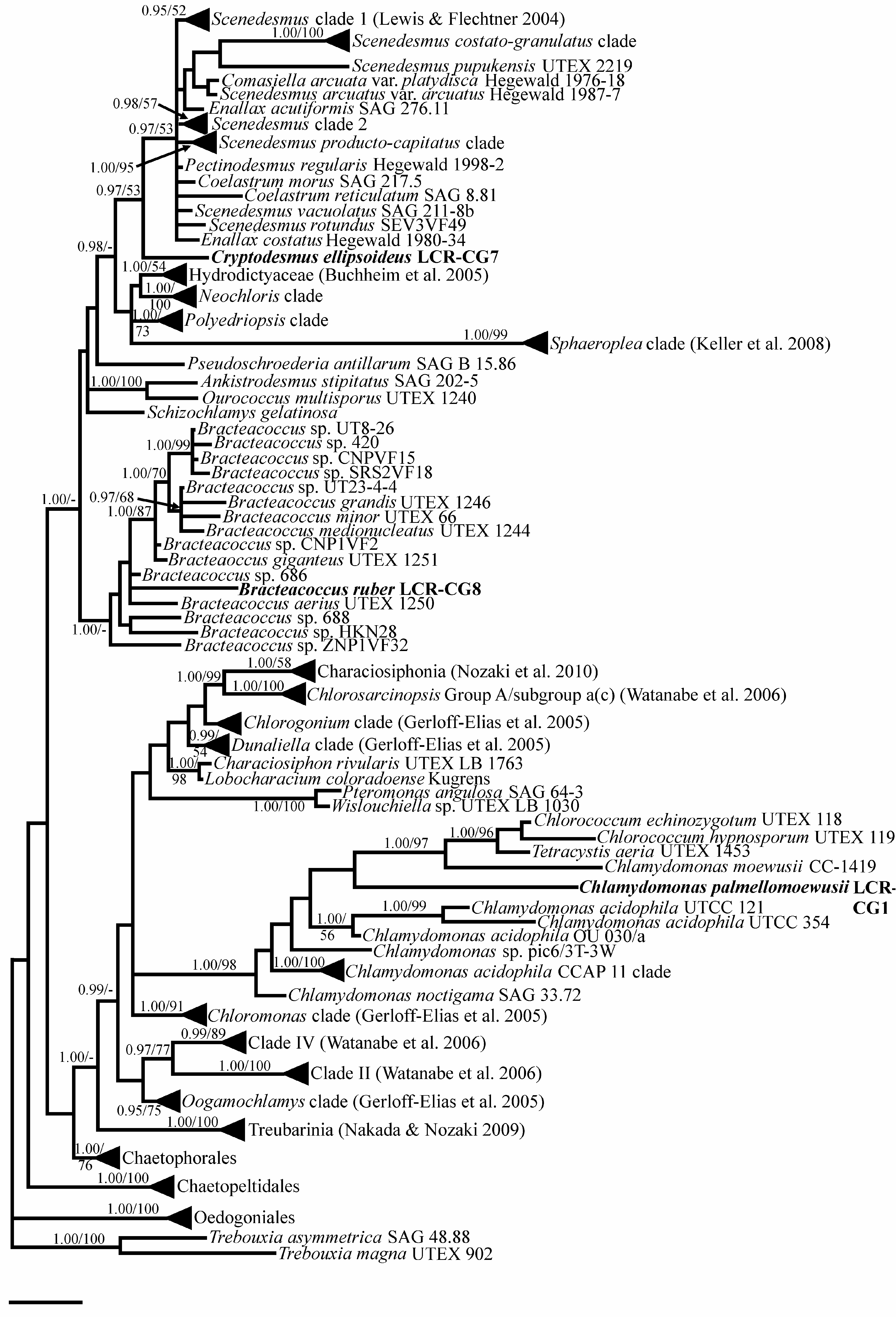
Cells ellipsoidal to slightly irregular, 6.2–9.2 µm long, 4.6–8.5 µm wide, single or in groups. Cell wall appearing smooth and thin in LM. Chloroplast cup-shaped, parietal, with single prominent pyrenoid traversed by one (–several) thylakoid membranes and encased in a segmented starch sheath of two (–several) parts ( Fig. 3E View FIGURE 3 ). Cell walls appear layered in TEM, especially in dividing cells, with irregular convoluted folds, especially in outer layers ( Fig. 3F View FIGURE 3 ). Reproduction by autospores, 2–4–8(–16) per sporangium ( Fig. 3B View FIGURE 3 ), often tightly packed; remnant mother cell walls frequently seen among cultured cells ( Fig. 3C View FIGURE 3 ). Very small protruberances are visible on the cell wall by TEM ( Fig 3E View FIGURE 3 ). DNA sequence data for the 18S gene suggest that this new genus represents a sister lineage to the clade formed by the genera Acutodesmus , Scenedesmus , Desmodesmus , Neodesmus , Pectinodesmus , Comasiella , and Coelastrum ( Fig. 7 View FIGURE 7 ).
Habitat:— Alpine herbfield soil.
Distribution:— New Zealand.
Etymology:— Referring to the relationship of this species with morphologically distinctive members of the genus Scenedesmus .
Observations:— Sequence data for rbc L were not obtained; amplified bands were 1 Kb larger than the standard size for the fragment, indicating the presence of a large intron and impeding sequencing.
Of particular interest, given the phylogenetic position of Cryptodesmus , are morphological features that might be homologous to more developed forms in Scenedesmus -like species (wall ornamentation, spines, coenobial colonies). A starch sheath surrounding the pyrenoid is found in Scenedesmus , but the morphology of the sheath and penetrating thylakoid described here is also very similar to that of Heterochlorella luteoviridis Neustupa et al. 2009 , a species placed unambiguously in the Watanabea clade, Trebouxiophyceae. It is also similar to that of Stichococcus jenerensis Neustupa et al. 2007, placed in the Prasiola clade, also in the Trebouxiophyceae. Pyrenoid morphology is therefore unhelpful. Scenedesmus species generally possess ornamented cell walls, and the walls of Cryptodesmus display an unusual layered structure, with slight evidence of thickenings resembling the processes sometimes found in Scenedesmus ( Fig. 3E View FIGURE 3 ). It is possible that these features share a common evolutionary origin. The dividing cells of Cryptodesmus are no more reminiscent of coenobial division than those of many other coccoid genera.
At least two globose–coccoid forms of Scenedesmus ( S. rotundus Lewis & Flechtner 2004 and Hylodesmus singaporensis Eliás et al. 2010 are already known ( Fig. 7 View FIGURE 7 )). Perhaps tellingly, these species with greatly reduced ornamentation are not found in fully aquatic habitats: S. rotundus was isolated from microbiotic crusts, and H. singaporensis from rotting wood. It is therefore possible that desiccation tolerance, and perhaps a reduced risk of predation, leads to the lack of the more spectacular ornamentations typical of Scenedesmus . The inclusion of two species of Coelastrum in the Scenedesmus clade, separated from Cryptodesmus , suggests that evolution of the features typical of Scenedesmus postdates the divergence between Scenedesmus and Cryptodesmus , and this may be resolved further if more diversity within a Cryptodesmus clade can be discovered.
Other species exist with similar internal structure, such as Pseudochlorococcum polymorphum ( Archibald 1970) , but these lack the external irregularities. In coccoids with external irregularities, such as Glochiococcus aciculiferus ( Silva 1996), Pseudotrochiscia ( Vinatzer 1975) , and Trochisciopsis ( Vinatzer 1975) , the protrusions are far more pronounced than in Cryptodesmus .
Cultures:— LCR-CG7.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |