
Cryptocellus leleupi Cooreman, 1976
|
publication ID |
https://doi.org/ 10.1206/3976.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03FC87DD-317D-0C5A-FE2B-B143FC7C4F05 |
|
treatment provided by |
Carolina |
|
scientific name |
Cryptocellus leleupi Cooreman, 1976 |
| status |
|
Cryptocellus leleupi Cooreman, 1976
Figures 1 View FIGURE 1 , 15 View FIGURE 15 , 16 View FIGURE 16 , 19C View FIGURE 19 ; table 1
Cryptocellus leleupi Cooreman, 1976: 27–50 , figs. 1–18; Platnick and Paz, 1979: 1 [considered nomen dubium]; Platnick, 2002: 386; Harvey, 2003: 179; Platnick and García, 2008: 145; Botero-Trujillo and Pérez, 2009: 57; Botero-Trujillo and Valdez-Mondragón, 2016: 330, 334, fig. 55; Botero-Trujillo and Flórez, 2017: 490.
Cryptocellus cf. leleupi: Fernández and Giribet, 2015: 4 .
TYPE MATERIAL: Three juveniles [1 protonymph holotype?, 1 deutonymph, 1 larva paratypes] ( IRSNB), ECUADOR: Oriente: Río Negro [01°24′32″S 78°11′28″W], under stones in transition forest, 1600 m (E.C.27), iv.1965, J. and N. Leleup; 1 juvenile [paratype?] ( IRSNB), same data except: forêt tropical à arehido, humus, 750 m (E.C.19), iv.1965, J. and N. Leleup. Photos examined GoogleMaps .
According to Cooreman (1976: 47), the type series of C. leleupi comprised four juveniles, but only three were found in the vial of specimens listed as types. A second vial containing another juvenile, with slightly different collection data from the original description, is assumed to be the fourth type specimen.
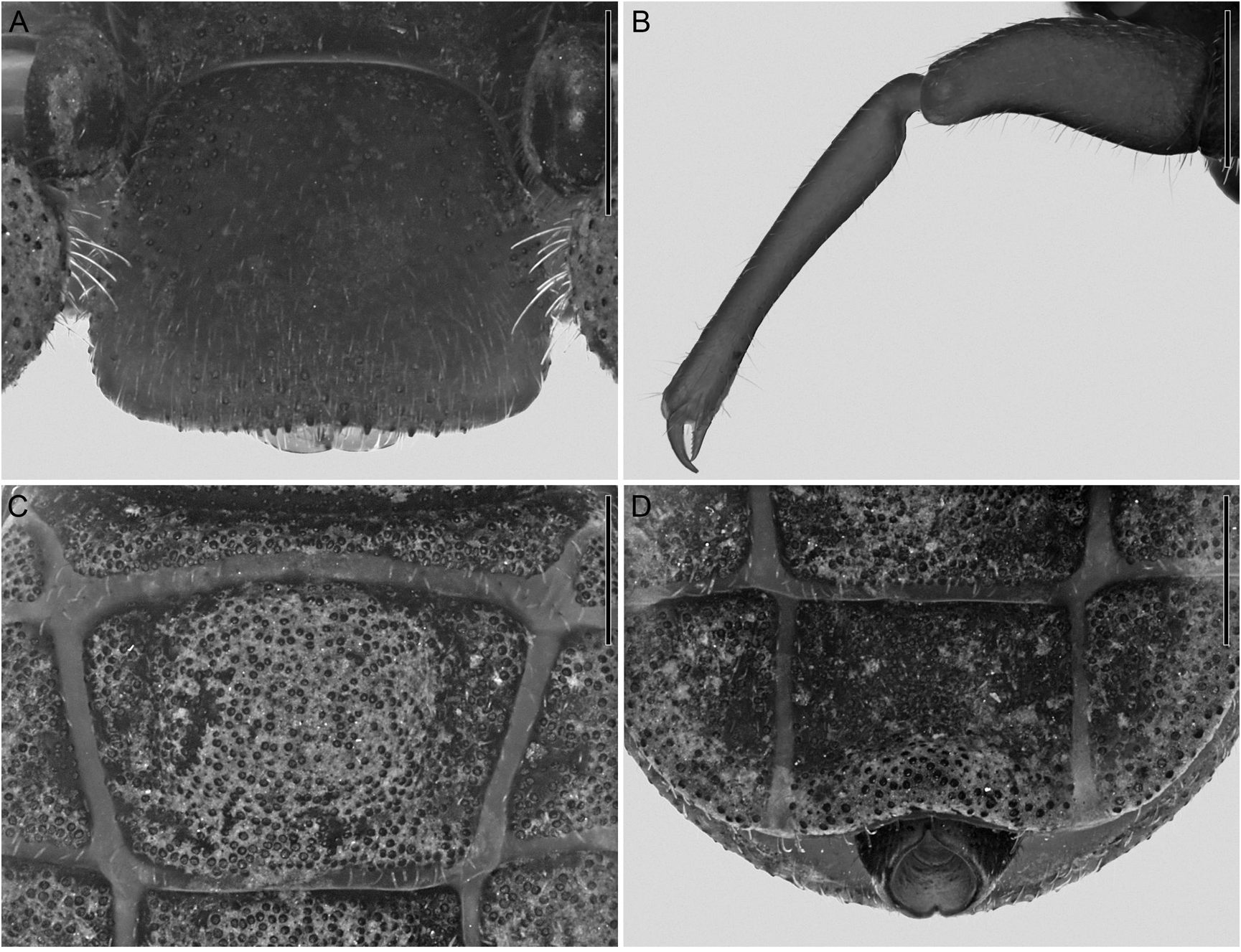
DIAGNOSIS: Cryptocellus leleupi closely resembles C. chiruisla and C. guaviarensis , with which it shares several morphological characters of the male: a pronounced median elevation and distinct, granular triangular surface of the cucullus; a wide movable finger of the chelicera; a well-developed ventral notch, ventral subbasal protrusion, and proventral distal expansion of the tibia of leg I; the metatarsus of leg I compressed prolaterally in proximal two thirds, with a distinct distal expansion ventrally; and a tuft of long setae ventrodistally on the metatarsus of leg III. The male of C. leleupi is most similar to that of C. chiruisla in the tibia of leg I, notably the subbasal protrusion situated proventrally in the proximal half of the segment, and the short ventral notch, visible in dorsal and prolateral aspects. The two species also resemble each other in the subapically truncated retrolateral margin of the cheliceral movable finger and in the similar shape of the fixed process of the male copulatory apparatus. The male of C. leleupi differs from the males of C. chiruisla and C. guaviarensis in the absence of a retroventral distal expansion of the tibia of leg I that, although small, is present in the other two species and in the anterior triangular surface of the cucullus, which is slightly concave in C. leleupi , but planar in the other species. Additionally, the posterodorsal margin of the basal segment of the pygidium is notched in C. leleupi and C. chiruisla . This notch is deep, with the ventral margin distinctly notched, in the female of C. leleupi (fig. 16D) (both are shallow in the male); in contrast, the pygidium is more shallowly notched dorsally, whereas the ventral margin is entire, in the female (and male) of C. chiruisla . Furthermore, the posterior genital lip is acute in the female of C. leleupi (fig. 19C) but trilobate in the female of C. chiruisla , and the spermathecal morphology also differs slightly in the two species.
MALE: The only male that has hitherto been located (UFMG 9099) is currently mislaid (see Remarks).
DESCRIPTION OF FEMALE: Based on the female from Estación Biológica Jatun Sacha (MACN Ar 37274).
Measurements: Total length, 4.26 mm (table 1).
Coloration: Soma and appendages predominantly reddish brown; pedipalps yellowish red. Carapace dorsolateral translucent areas yellow. Cheliceral manus yellow; fingers and finger dentition reddish.
Setation: Soma and appendages covered with fine, translucent, bristlelike setae (figs. 15, 16).
Tegument surface macrosculpture: Tegument without cuticular pits. Carapace densely covered with conspicuous, round, iridescent granules, sparsely granular anteriorly (fig. 15A). Cucullus sparsely granular, granulation similar to carapace, with some larger granules ventrally (fig. 16A). Coxosternal region predominantly smooth, intercoxal margins finely granular (fig. 15B). Opisthosoma dorsal and ventral surfaces densely granular (fig. 15C, D), similar to carapace. Pedipalp predominantly smooth (fig. 16B), retrolateral surfaces of trochanter 1, trochanter 2, and base of femur finely granular. Legs with granulation similar to carapace, but more sparse.
Carapace: Carapace trapezoidal, approximately as long as wide (table 1), broadest between coxae of legs II and III; lateral margins curved, converging anteriorly (fig. 15A); anterior margin linear in dorsal aspect, sublinear in frontal aspect; posterior margin procurved; median longitudinal sulcus shallow, partial; paired lateral depressions present near lateral margins, adjacent to coxae of legs II; dorsolateral translucent areas well defined, visible in dorsal and lateral aspects, glabrous and slightly convex, aligned with intersection between coxae of legs I and II.
Cucullus: Cucullus trapezoidal, wider than long (table 1); lateral margins diverging ventrally (fig. 16A); ventral margin predominantly linear in anterior aspect, shallowly concave in ventral aspect.
Chelicerae: Movable finger longer than fixed finger, not widened; mucron sharp; tooth row comprising 10 or 11 small teeth. Fixed finger tooth row comprising five small, sharp teeth and one markedly larger distal tooth.
Coxosternal region: Tritosternum moderately small, tuberculate, not abutting coxae of legs I (fig. 15B); coxae of legs II–IV abutting one another medially along entire length; coxae of legs II, anterior and posterior margins not perpendicular to median axis, forming angle medially; coxae of legs II–IV progressively decreasing in length (table 1); sutures between coxae of legs II and III slightly longer than suture between coxae of legs IV.
Opisthosoma: Opisthosoma oval, longer than wide (fig. 15C, D, table 1), broadest at tergite XII. Median sclerite of tergite X slitlike, trapezoidal; of tergites XI–XIII each with paired, shallow submedian depressions, lateral margins converging posteriorly on XI, subparallel on XII and XIII; of tergites XI–XIII wider than long (table 1); of tergite XI with raised surface medially (fig. 16C); of tergite XIII, lateral margins forming right angle with posterior margin, posterior corners not protruding laterally. Sternites XI–XIII each with paired submedian depressions similar to tergites (fig. 15D). Pygidium, basal segment posterior margin with distinct notch dorsally and ventrally (fig. 16D).
Pedipalps: Femur robust (table 1), dorsal surface convex. Tibia longer than femur (table 1), entirely linear (fig. 16B), moderately swollen proximally, rest of segment noticeably narrower; elevated oval tubercles absent. Movable finger longer and more robust than fixed finger.
Legs: Leg II longest; I–IV similar in width, no segments swollen (table 1). Legs I and II femora, dorsal and ventral surfaces each with moderate projection on proximal end, protecting articulation with trochanter. Legs unmodified.
Spermathecae: Spermathecae heavily sclerotized, wider than deep (fig. 19C), separated from one another by approximately one seventh of spermathecal width; internal cavities markedly convoluted, pattern of convolution bilaterally asymmetrical. Anterior and posterior genital lips subtriangular (fig. 19C); anterior lip apex acuminate, posterior lip apex acute. Pair of small papillate vesicles situated submedially on posterior surface, aligned with base of spermathecae (fig. 19C).
DISTRIBUTION: Cryptocellus leleupi has been recorded from the Jatun Sacha Biological Station and the Río Negro, respectively situated in the Napo and Tungurahua provinces of Ecuador (fig. 1) .
REMARKS: Cryptocellus leleupi was described from a juvenile specimen which, based on the number of tarsomeres on the walking legs (I:1, II:4, III:3, IV:2), can be recognized as a protonymph. The type series consists entirely of juveniles, thus failing to provide information concerning the adult morphology necessary for species diagnosis and was therefore considered a nomen dubium by Platnick and Paz (1979).
No other published records of ricinuleids occur in the immediate proximity of the type locality of C. leleupi , Río Negro, situated in the eastern foothills of the Ecuadorian Andes. However, a juvenile identified as Cryptocellus cf. leleupi , from the Jatun Sacha Biological Station, situated ca. 73 km northeast of the type locality, was included in a molecular study by Fernández and Giribet (2015). The taxonomic identification of the cytochrome oxidase c subunit I DNA sequence (GenBank accession KR180410 View Materials ) is listed as C. leleupi although the voucher (IZ-130032) is listed as Cryptocellus sp. in MCZbase, the database of the Zoological Collections (https://mczbase.mcz.harvard.edu/SpecimenSearch.cfm).
A male and a female from the Jatun Sacha Biological Station were examined during the present investigation and confirmed to differ from all other ricinuleid species. Given the relative proximity of this station to the type locality of C. leleupi , it seems reasonable to consider these specimens to be conspecific.
The male (UFMG 9099) was examined in December 2017, when the species diagnosis was prepared. Soon after, the vial with the specimen was mislaid in UFMG and, despite extensive searches, has not been located, preventing a more detailed and thorough documentation of the male morphology. Nevertheless, the diagnosis presented herein is sufficient to permit recognition of the species. The structures noted in the diagnosis have been well documented in the description of the two other Cryptocellus species to which C. leleupi is compared (Botero- Trujillo and Flórez, 2017).
Although knowledge of the adult morphology of C. leleupi remains incomplete, it is now possible to recognize this species. Therefore, C. leleupi is no longer considered a nomen dubium.
MATERIAL EXAMINED: ECUADOR: Napo: Cantón Tena, Parroquia Puerto Napo, Estación Biológica Jatun Sacha , 01°03′57.5″S 77°37′0.2″W, 410 m, 1–5.xii.2009, C. Grismado and F. Labarque ( Oonopidae Planetary Biodiversity Inventory Expedition ), humid forest, leaf litter, 1 ♀ ( MACN Ar 37274) GoogleMaps , same data, except: A.J. Santos, 1 ♂ GoogleMaps ( UFMG 9099 View Materials ) .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cryptocellus leleupi Cooreman, 1976
| Botero-Trujillo, Ricardo, Carvalho, Leonardo S., Florez D., Eduardo & Prendini, Lorenzo 2021 |
Cryptocellus leleupi
| Harvey, M. S. 2003: 179 |
| Platnick, N. I. 2002: 386 |
| Cooreman, J. 1976: 50 |