
Coelorinchus mascarenus, Prokofiev & Iwamoto, 2020
|
publication ID |
https://doi.org/ 10.5281/zenodo.11105838 |
|
DOI |
https://doi.org/10.5281/zenodo.11105878 |
|
persistent identifier |
https://treatment.plazi.org/id/3A3FD645-BD21-4B49-A235-F6E9B73AE672 |
|
treatment provided by |
Felipe |
|
scientific name |
Coelorinchus mascarenus |
| status |
sp. nov. |
Coelorinchus mascarenus new species
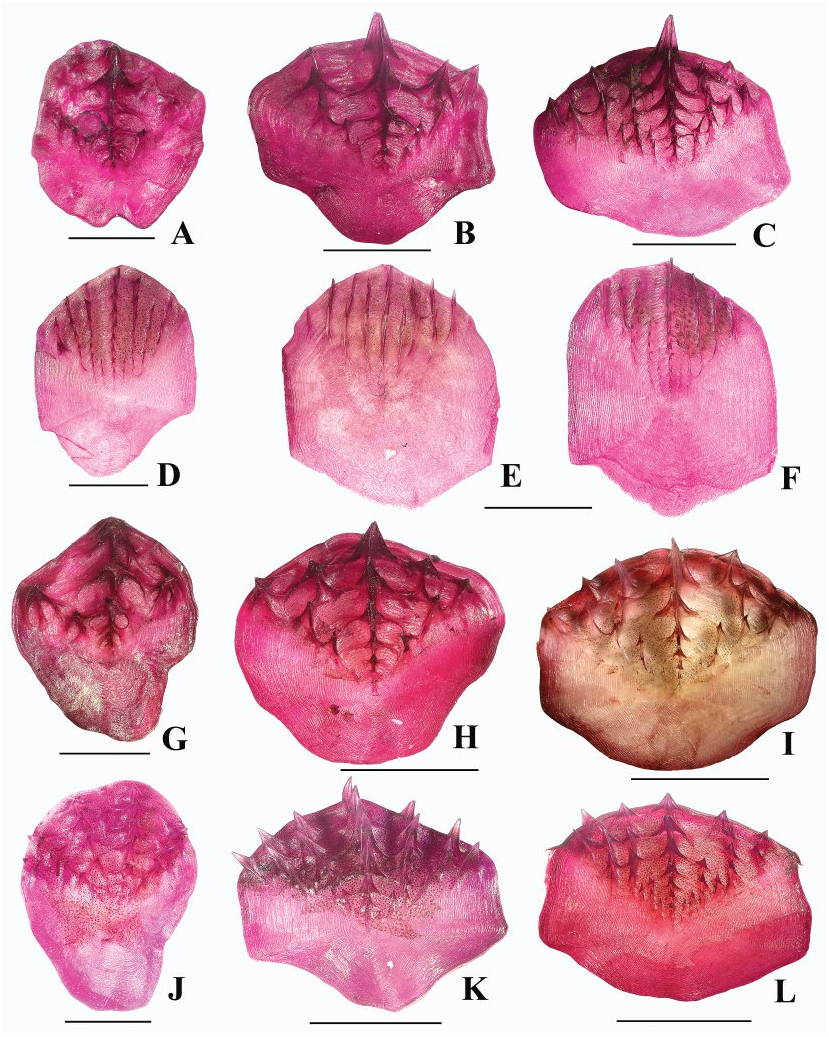
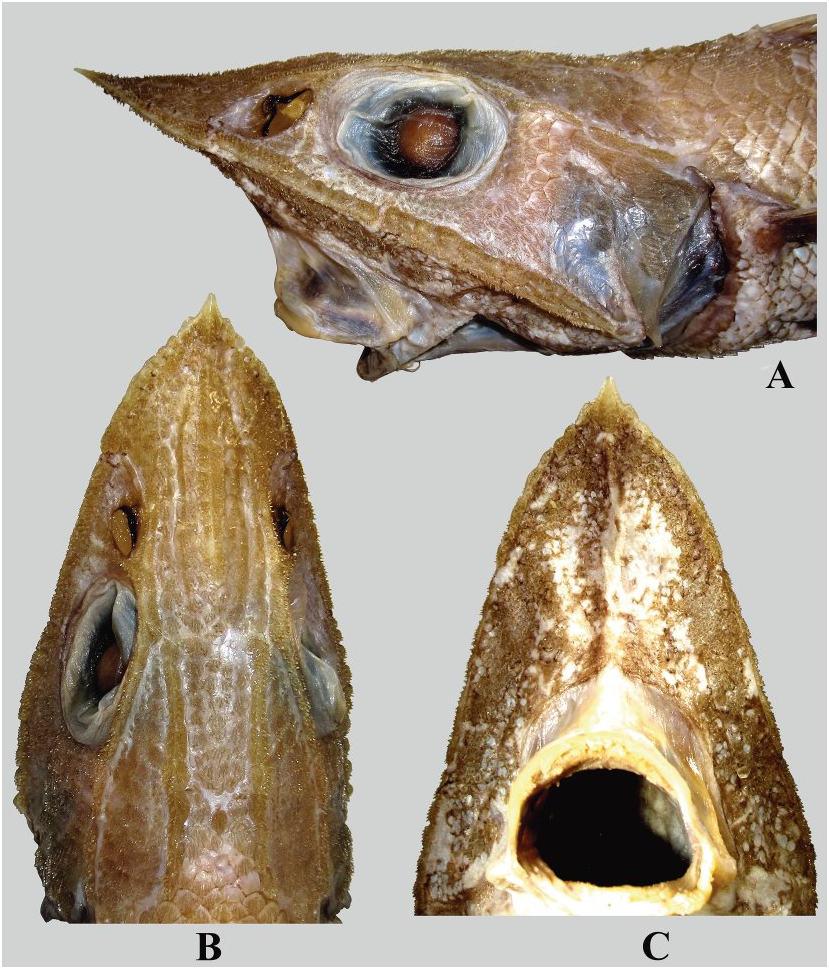
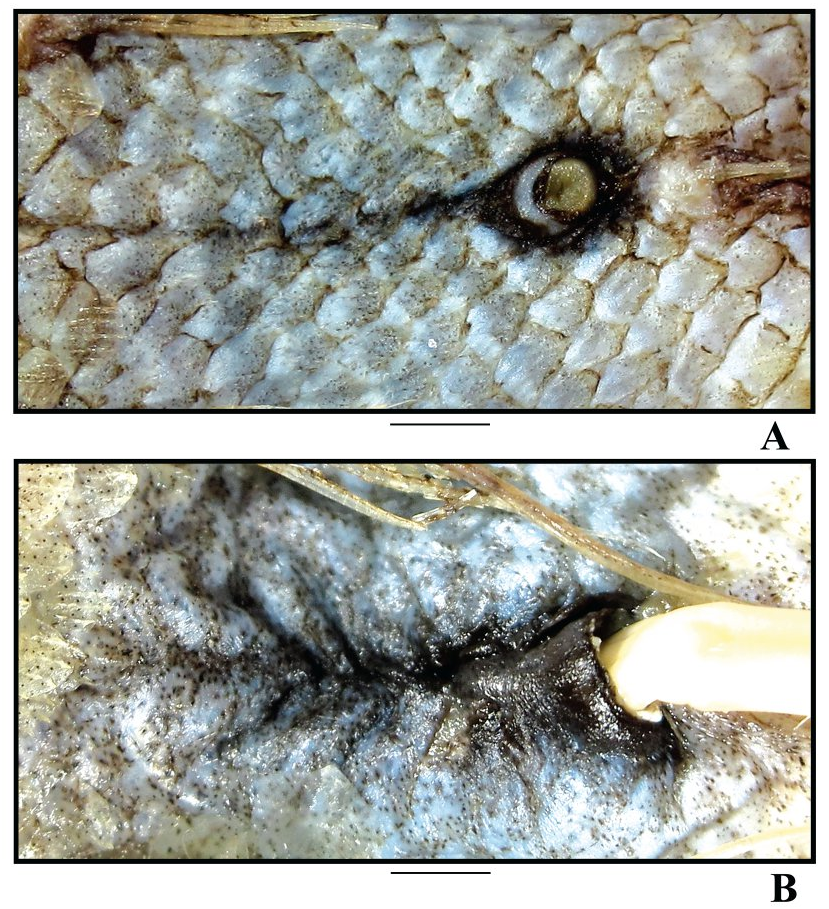
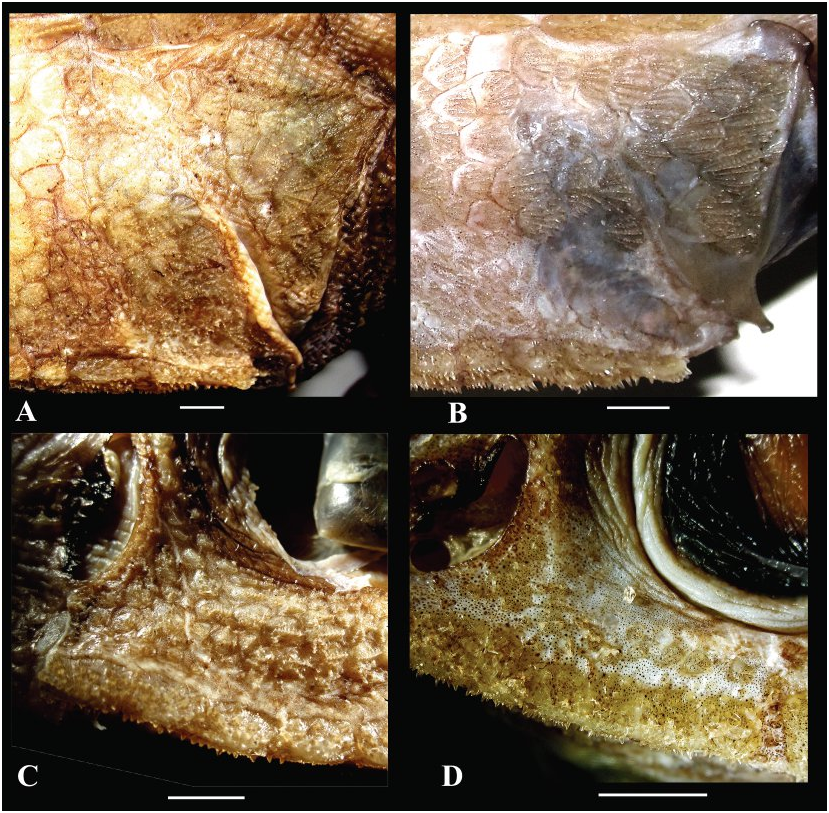
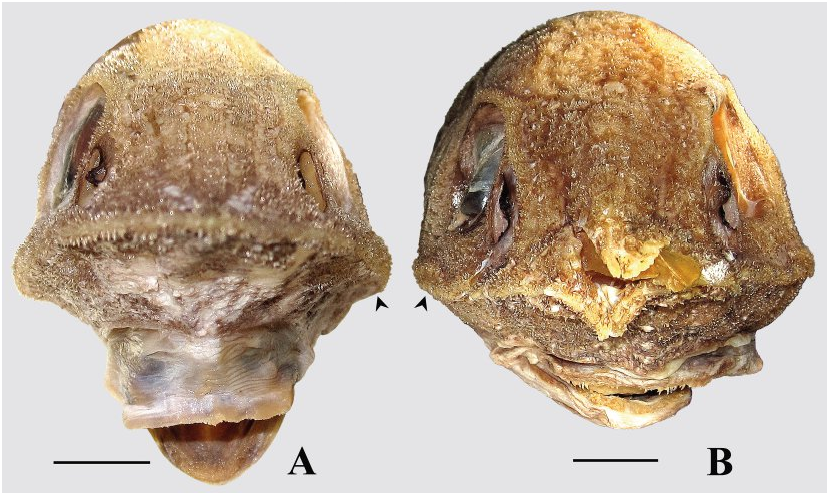
Figures 2B View FIGURE , 5D–F View FIGURE , 10 View FIGURE , 11A View FIGURE , 12B, 12D View FIGURE , 13A View FIGURE .
Coelorinchus yurii (partim.) Iwamoto, Golani, Baranes and Goren, 2006:434 (all paratype specimens; Saya de Malha Bank, Mascarene Ridge, 800–1130 m).
Holotype. — IOM M.011–007 (91 mm HL, 297+ mm TL), Mascarene Ridge , off Saya de Malha Bank, 8°32´S, 59°41´E, 960–1130 m, R / V Vityaz-II, station 2820, 29-m otter-trawl, 10 Jan. 1989. GoogleMaps
Paratypes. — IOM M. 012–019 (1, 93 mm HL, 260+ mm TL) and CAS 66427 About CAS (2, 84.7–94. 3 mm HL, 275+– 313 mm TL), same data as for holotype GoogleMaps . CAS 223466 About CAS (3, 72–76. 5 mm HL, 245– 290 mm TL) and IOM M.013–000 (7, 72–95 mm HL, 250+–330+ mm TL), Mascarene Ridge , Saya de Malha Bank, 9°32.7´S, 60°02´E, 800 m, FRV Fiolent, cruise 7(9), trawl 156, 26 Sept. 1977 GoogleMaps .
NOTE.— We could not find the 84.7-mm HL paratype of C. yurii ( FRV Fiolent, cruise 7(9), trawl 156A) listed by Iwamoto et al. (2006) for the current study; thus, its identification remains uncertain.
DIAGNOSIS.— A species of Coelorinchus acanthiger –group with light organ externally represented by a short cleft anterior to anal surround, its length 3.0–9.9 % HL; snout length 39–43 % HL, length medial element of terminal scute 6+–8.8 % HL; preoral region partially to almost completely scaled at 72 mm HL or greater, nasal fossa scaleless, triangular area between nasal fossa, orbit, and suborbital ridge sparsely scaled; scales over interorbital space with irregularly disposed spinules, not forming clear keel-like rows; scales on body loosely attached, bearing spinules with buttresses weak or absent; 3.0–4.5 and 4.0–5.0 scales between midbase of first dorsal fin and between origin of second dorsal fin and lateral line, respectively; body color light, fins darker, lips and gums pale to dusky.
DESCRIPTION.— General features of fish seen in Figs. 2B View FIGURE , 10 View FIGURE . Counts: first dorsal-fin rays ii + 8; pectoral-fin rays i + 16–18 [i + 17]; pelvic-fin rays 7; gill-rakers (inner) on 1 st arch 7 (8, as exception) [7]; gill-rakers on 2 nd arch 5–6 (usually 5) (outer) / 7 (as exception, 8) (inner) [5 / 7]; transverse scale rows below origin of first dorsal fin 3.5–4.5 [4.5]; ditto, below midbase of first dorsal fin 3.0–4.5 [3.5]; ditto, below origin of second dorsal fin 4.0–5.0 (usually 4.5) [4.0]; ditto, between origin of anal fin and lateral line 10–15 [15]; lateral-line scales before origin of second dorsal fin 14–15 [14]; pyloric caeca 6–11 (n = 5).
Measurements shown in Table 2 View TABLE . Width of body across pectoral bases 74.3–86.4 % of greatest body depth (at dorsal-fin origin), 1.1–1.3 times smaller than greatest width of head (at preopercles). Head c. 2.8–3.5 (usually 3.3 or more) times in TL. Snout long, 2.4–2.6 times in HL, 1.1–1.3 times greater than postorbital length of head. Snout acute, slightly upturned (usually) to almost horizontal in lateral view, with dorsal contour very weakly concave ( Fig. 10A View FIGURE ); distinctly convex at sides in dorsal view, with sides much more strongly convergent toward the tip in anterior third of snout length ( Figs. 10B, 10C View FIGURE ). Snout tipped with moderately long, somewhat diamond-shaped terminal scute, which lateral elements form conspicuous “shoulders” aside medial element. Anterolateral margins of snout not completely supported by bone. Orbit elliptical, 3.7–4.3 times in HL, 1.5–1.7(1.8) times in snout. Suborbital shelf angulated ( Fig. 13A View FIGURE ); shelf depth 1.7–2.2 times in suborbital depth. Lateral nasal ridge 3.2–3.3 times shorter than suborbital ridge. Mouth comparably large, posterior tip of maxilla extending below posterior third to posterior quarter of orbit, rictus below middle of orbit. Preopercle inclined backward at about 60–70º, with moderately long posteroventral lobe more or less angularly rounded at tip. Subopercle terminates ventrally in slender tip that extends beyond preopercle. Chin barbel short, slender, 2.8–3.6 (usually 3.0–3.4) times in orbit. Free neuromasts on snout, along head ridges and on underside of head prominent, vary in color from whitish to blackish (always more or less infuscated on top and underside of snout), surrounded by isolated or scattered black hair-like papillae, more densely aggregated on preoral portion of underside of head below suborbital ridge and in front of upper jaw, few or absent in nasal fossa, behind mouth and on top of head above and behind orbits. Inner side of gill opening with or without fleshy protrusion in its lower half.
Anus close to anal-fin origin. Light organ externally evident as a short cleft, not forming scaleless black fossa, 3.1–10.4 times in pelvic-anus distance, its anterior end often poorly discernible, surrounded by scale pockets bearing dense aggregations of large black melanophores on silvery background ( Fig. 11A View FIGURE ). Area on belly occupied by such melanophores varies in size from specimen to specimen, at maximal development it covers scale pockets from anus anteriad to a line between pelvic-fin origins and up to three longitudinal rows of scales on each side laterally from midventral line.
Jaw teeth conical, pointed, arranged in bands of nearly uniform width; premaxillary teeth larger than these on dentary; outermost teeth of premaxilla weakly to not enlarged. Premaxillary tooth band short, 1.7–2.0 times in length of rictus; dentary tooth band reaching rictus or nearly so.
First dorsal-fin base 1.2–1.6 times shorter than the interdorsal space; second dorsal-fin spine ending in filament, which length approximately equal to postrostral length. Pectoral fin narrow based, rather short, falling to or slightly before anal-fin origin. Pelvic fins originate more or less behind the vertical of pectoral-fin origin (rarely on the same vertical). Outermost pectoral-fin ray filamentous and extending to or just behind anal-fin origin.
Squamation ( Figs. 5D–F View FIGURE , 12B, 12D View FIGURE ). Body scales deciduous, those on head and predorsal region more adherent. Scales large with spinules in parallel rows (parallel to slightly divergent on scales in predorsal region). Spinules on scales below first dorsal-fin base arranged in 7–10 rows, some rows often incomplete; those on scales below beginning of second dorsal fin arranged in 9–12 rows, some often incomplete; middle row of spinules somewhat enlarged, giving appearance of horizontal striations on body surface; spinules in rows gradually increasing in length posteriad (similarly expressed on flanks and in predorsal region). Lateral buttresses of spinules weakly developed to almost absent, never joined with those of neighboring rows. Predorsal scales with spinules only slightly more raised and less depressed than on flank scales; spinules on predorsal scales arranged in 5–8 (usually 6 or 7) rows. Scales on top of head, cheeks, and gill cover with parallel to slightly diverging rows of spinules ( Fig. 12B View FIGURE ); scales on interorbital space irregularly disposed, not forming clear keel-like rows; area between the orbit, nasal fossa, and suborbital ridge sparsely scaled, with spinules on scales arranged in clusters, rarely forming a single short row on some scales. Narrowest portion of suborbital shelf bearing 1 (rarely 0, in smallest specimen examined) to 2 rows of scales between lower rim of orbit and upper edge of suborbital ridge ( Fig. 12D View FIGURE ); these scales often separated from each other by bare interspaces, none enlarged. Nasal fossa completely scaleless. Top of snout densely scaled. Underside of head usually densely scaled except on gular region, branchiostegal membranes, and small triangular area in front of premaxillary symphysis (rarely middle portion of preoral region largely scaleless); scales strongly deciduous (mostly missing) in central area of preoral region and behind mouth. Scales from underside of head with spinules arranged in 1–4 short widely separated rows (1–4 spinules per row). Scutes of head ridges strongly armed with numerous stout conical spinules with pointed tips; scutes of medial nasal ridge with radiating rows of conical spinules lacking buttresses, 9–10 (11) in number (including terminal scute). Supraoccipital and postoccipital scutes very small (commensurable with neighboring scales), coarsely spinulated.
Body color pale brownish without markings. Light-brown coloration formed by dense aggregations of minute brownish melanophores on epithelium covering exposed field of scales. When scales removed, scale pockets pure white with narrow dark-brown margins and sparse melanophore peppering in posterior third of scale pocket (more conspicuous below lateral line). Orbit not encircled by dark ring; anterior border of second naris blackish. Scaled parts of underside of head darker than dorsal and lateral surfaces of head (coloration formed by dense aggregation of brownish melanophores); gular region and branchiostegal membranes from paler than, to as dark as, preoral region; distal margin of branchiostegal membrane blackish in its upper half. Lips and gums pale to dusky, mouth cavity dusky, gustatory papillae vary in color from white to brownish or blackish; branchial cavity and peritoneum blackish; stomach pale. All fins more or less dusky to blackish (rays of second dorsal fin pale), scale pockets at bases of anal-fin rays much more densely peppered with melanophores than neighboring ones.
ETYMOLOGY.— Named from its distribution area, Mascarene Ridge.
REMARKS.— All paratypes of C. yurii ( Iwamoto et al., 2006:434) are referred to this species. Although it was formerly believed that differences between the holotype and paratypes can be attributed to size-related changes ( Iwamoto et al., 2006:438), some of these differences (presence or absence of external light organ, adherent vs. deciduous scales) are clearly not this case. Furthermore, the 81–85 mm HL ( IOM 013–000) paratypes are adults with mature gonads (including females 81 and 82 mm HL with ripe eggs about 0.5 mm in diameter). As C. mascarenus is very similar and apparently very closely related to C. braueri , it is probable that our specimens of C. mascarenus attain sizes close to maximum for this species. Both the holotype and the second known specimen of C. yurii described herein are much larger (154 and 156 mm HL) than the type series of C. mascarenus ; thus, it is not possible to compare these species with specimens of similar size. However, we have a graded size series of C. amirantensis , a species close to C. yurii , containing specimens of similar size to both C. mascarenus and C. yurii . Results of our analysis show that most differences between C. mascarenus and C. yurii are not correlated with growth in C. amirantensis or sometimes show opposed trends ( Table 3 View TABLE ).
COMPARISONS.— The new species is similar to C. braueri known from off South Africa and seamounts in the southwestern part of the Indian Ocean northward to about 25°S. The main difference between these species include the external manifestation of the light organ, narrow and cleft-like, lacking a dermal window in C. mascarenus , compared with oval black naked fossa in C. braueri ( Fig. 11 View FIGURE ). Furthermore, the new species has more deciduous scales and less attenuated snout than C. braueri , with shorter terminal scute (length 6+–8.8 vs. 9.5–12.2 % HL, n = 20), a smaller orbit (23–27, usually 25 % HL vs. 26–34, usually 28–32 % HL, n = 36, in C. braueri ), and on average fewer scales below midbase of first dorsal fin and lateral line (usually 3.5 vs. usually 4.5 in C. braueri , n = 36) and scales between origin of second dorsal fin and lateral line (usually 4.0–5.0 vs. usually 5.0–5. 5 in C. braueri , n = 36). For comparison with C. yurii , see Table 3 View TABLE .

| IOM |
Institute of Oceanology, Academy of Sciences |
| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |