
Coelogynopora kenichii, Volonterio & León, 2021
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.775.1557 |
|
publication LSID |
lsid:zoobank.org:pub:0F448B63-A29C-4F5B-995D-7C8A84089153 |
|
DOI |
https://doi.org/10.5281/zenodo.5604697 |
|
persistent identifier |
https://treatment.plazi.org/id/FC85AFB9-55C1-47C4-8A5F-AEA9FD3D3D22 |
|
taxon LSID |
lsid:zoobank.org:act:FC85AFB9-55C1-47C4-8A5F-AEA9FD3D3D22 |
|
treatment provided by |
Felipe |
|
scientific name |
Coelogynopora kenichii |
| status |
sp. nov. |
Coelogynopora kenichii sp. nov.
urn:lsid:zoobank.org:act:FC85AFB9-55C1-47C4-8A5F-AEA9FD3D3D22
Figs 1–6 View Fig View Fig View Fig View Fig View Fig 5 View Fig
Diagnosis
Coelogynopora kenichii sp. nov. is defined by the following combination of characters: sclerotised copulatory system consisting of a slender, ventrally curved stylet with a broad base, and three pairs of symmetrically arranged spines, the proximal ends of which are fused laterally to the base of the stylet. Distal ends of the three pairs of spines hooked, with apophyses at progressively longer distances from the tip. Accessory spines and solar organ absent.
Etymology
We dedicate this species to Prof. Dr Ken-Ichi Tajika for his highly relevant contributions to the systematics of Coelogynoporidae and other Proseriata .
Type material
Sixteen whole mounts and two series of sagittal and transversal sections.
Material examined
Holotype CHILE • Whole mount, stained with carmine and fast green, mounted in Canada balsam; coast of the Magellan Strait, Magellan Province; 53.016667° S, 70.816667° W; 6 Jan. 2005; R. Ponce de León leg.; RMNH.VER.19980.a. GoogleMaps
Paratypes CHILE • 16 whole mounts, 2 series; same collection data as for holotype. RMNH.VER.19980.b to RMNH.VER.19980.k (10 whole mounts, stained with carmine and fast green, mounted in Canada balsam), RMNH.VER.19980.l (a series of sagittal sections stained with hematoxylin-eosin), RMNH. VER.19980.m (a series of transversal sections in four slides, stained with hematoxylin-eosin), RMNH.VER.19980.n (two whole mounts in one slide, cleared and mounted in lactophenol), RMNH. VER.19980.o (a whole mount, cleared and mounted in lactophenol) and RMNH.VER.19980.p (three whole mounts in one slide, cleared and mounted in lactophenol) GoogleMaps .
Description
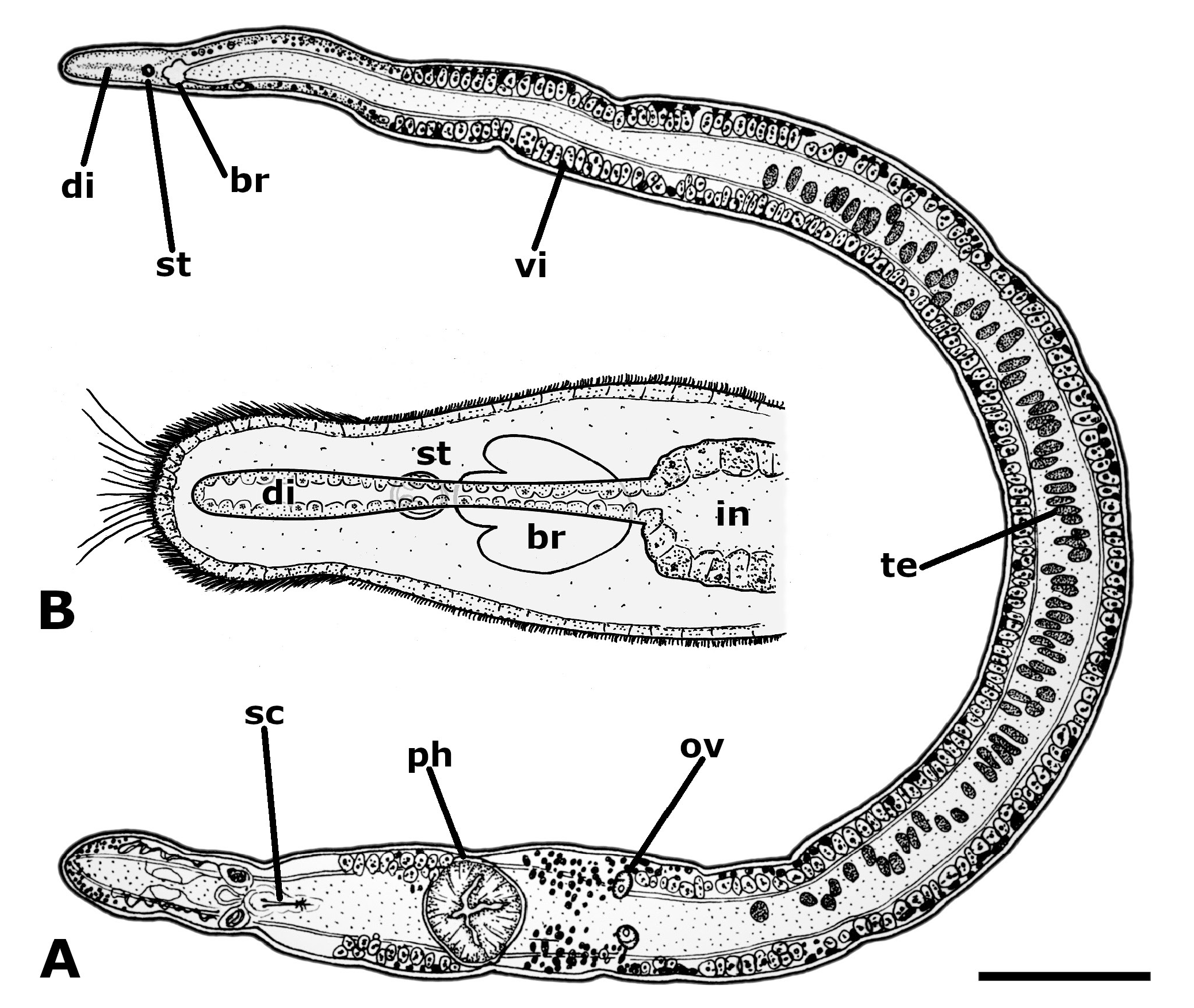
BODY SIZE. Live, adult animals filiform, 5.24–8.82 mm long (6.47, 1.610, 4). Fixed specimens 2.57– 5.72 mm long (3.92, 0.981, 11) and 125–286 µm (198, 50.3, 10) wide at the level of the ovaries ( Fig. 1A View Fig ).
SENSORY ORGANS AND GLANDS. Anterior end of the body provided with short sensory bristles and a few long tactile hairs ( Fig. 1B View Fig ). Two types of gland cells open through the anterior epidermis: one that stains green with fast green, and another one, smaller, that does not. Encapsulated brain, anteriorly trilobate, at about 4.4% from the anterior end of the body ( Fig. 1B View Fig ); 54–82 µm (64, 8.8, 9) long, and 40–67 µm (50, 9.4, 9) maximum width. Statocyst with two pairs of statoliths at about 3.2% from the anterior end ( Fig. 1B View Fig ), 11–16 µm (14, 1.7, 7) in diameter. At least three types of parenchymal gland cells, with different affinities for the stains used, open through the epidermis of the general body surface. An aggregation of gland cells is also present in the posterior end of the body. Paracnids absent.
DIGESTIVE SYSTEM. Intestine with an anterior, hollow pre-cerebral diverticulum–“ Kopfdarm ” or “ chorda intestinalis ” sensu Ax (1957) and Ax & Ax (1969) – that arises at the level of the posterior part of the brain, continues anteriorly as a narrow tube, dorsally to the brain and the statocyst, and widens anteriorly to the latter, extending almost to the anterior end of the animal ( Fig. 1B View Fig ). The diverticulum is lined with a thin wall and has a wide lumen except in the region directly above the brain and statocyst; intestinal contents were seen in transversal sections throughout its length. The pharynx, directed ventrally, is at about 75% from the anterior end and has numerous anterior and posterior associated glands ( Fig. 1A View Fig ).
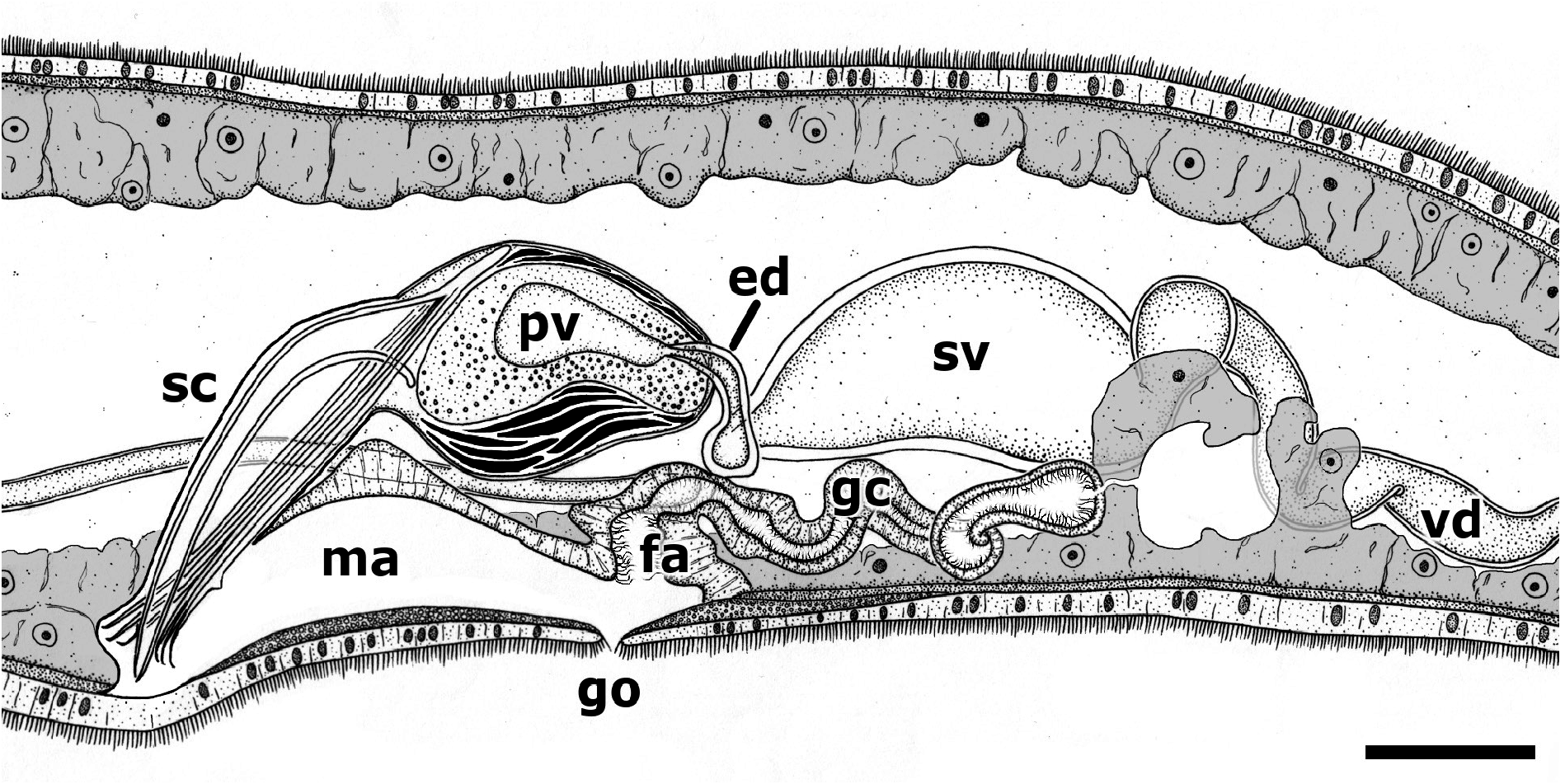
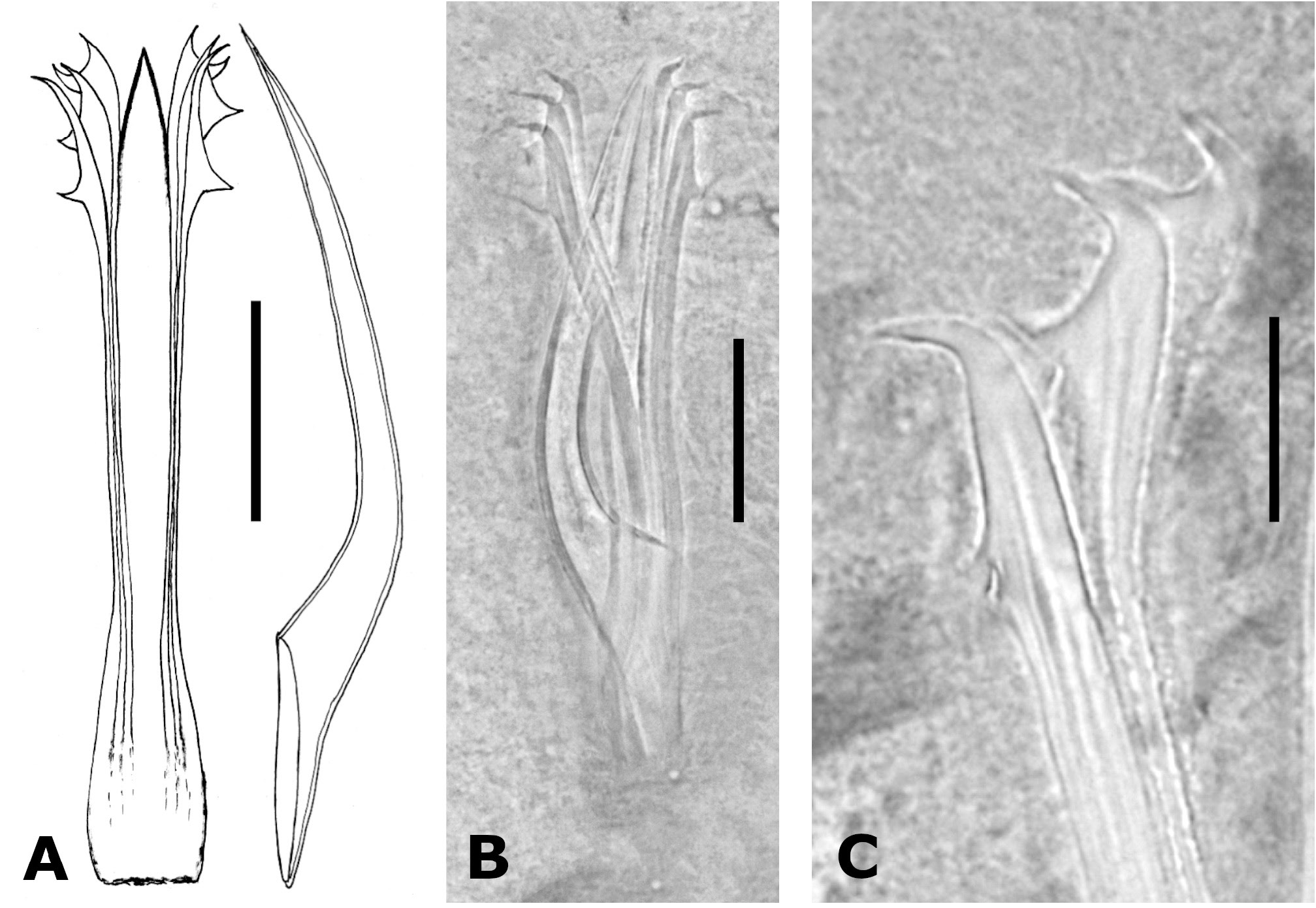
MALE REPRODUCTIVE SYSTEM. An irregular median row of 41–85 (55, 13.4, 10) testicular follicles extends from 26% to 66% from the anterior end ( Fig. 1A View Fig ). Each follicle is 42–63 µm (50, 5.3, 24) long, 15–33 µm (21, 5.2, 24) wide. Two long vasa deferentia extend posteriorly almost to the end of the body, turn 180 degrees, and run anteriorly following a sinuous course, opening into the proximal end of each of the two seminal vesicles ( Fig. 2 View Fig ). The seminal vesicles open into a common ejaculatory duct that runs dorsally and enters the prostatic vesicle ( Figs 2–3 View Fig View Fig ). Several prostatic glands surround the ejaculatory duct and the proximal part of the prostatic vesicle, opening into the latter. Oblique muscle fibres surround the prostatic vesicle, giving a continuous layer that is thicker ventrally. The sclerotised copulatory system lies anterior to the gonopore, is directed downwards and forwards, and opens into the anterior end of the genital atrium ( Fig. 3 View Fig ). It consists of a stylet and 3 pairs of symmetrically arranged spines ( Fig. 4 View Fig ). The stylet is slender, ventrally curved, and is provided with a broad base ( Fig. 4A View Fig ); it is 77–97 µm long (90, 6.4, 10) and 16–24 µm (20, 2.7, 10) in basal width. The three pairs of spines are 75–90 µm (84, 4.6, 10), 78–93 µm (88, 5.0, 10) and 81–97 µm (91, 5.1, 10) in length; the smallest are the farthest from the stylet. The proximal ends of all three pairs of spines are fused laterally to the base of the stylet ( Fig. 4A–B View Fig ); their distal ends are hooked, and their apophyses are placed at progressively longer distances from the tip ( Fig. 4C View Fig ).
FEMALE REPRODUCTIVE SYSTEM. The vitellaria consist of many follicles in two rows that are lateral to the intestinal sac; they extend from well behind the brain to about the level of the anterior part of the genital atrium ( Fig. 1A View Fig ). Two elliptical ovaries are approximately halfway between the level of the more posterior testes and the anterior end of the pharynx ( Fig. 1A View Fig ); they are 36–75 µm (54, 12.1, 11) long and 24–37 µm (31, 4.0, 11) wide. Two ciliated ovovitelline ducts run posteriorly and open laterally into the ciliated, caudal end of the genital atrium, just behind the gonopore. A winding, narrow genital canal leads from the posterior atrial wall to the intestinal wall; it opens into the intestinal lumen, giving a genito-intestinal connection ( Fig. 3 View Fig ). The canal is thick-walled and lined with ciliated epithelium. The distalmost portions of the ovovitelline ducts, the genital canal and the caudal atrial wall are surrounded by glands.
The genital pore is at about 80% from the anterior end, at the level of the posterior end of the prostatic vesicle ( Figs 1A View Fig , 3 View Fig ); it does not have an obvious sphincter.
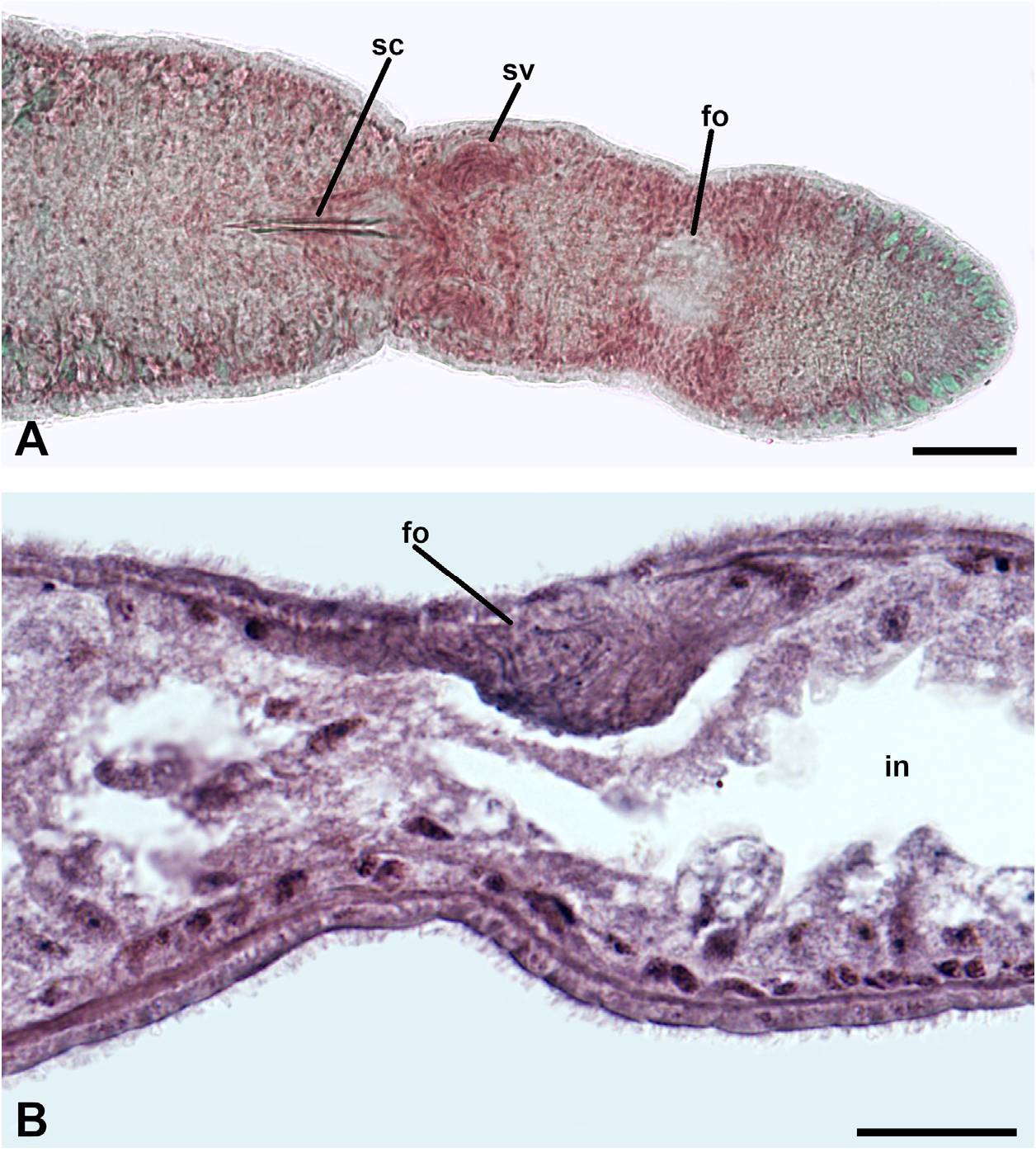
FIBROUS ORGAN. An unknown structure was observed in every specimen studied, behind the reproductive complex and at the level of a posterior constriction of the body. In whole mounts, it appears as a welldefined, clear area ( Fig. 5A View Fig 5 ); in histological sections, it is evident as a dorsal, fibrous mass between the body wall and the intestine that extends anteriorly for a short distance ( Fig. 5B View Fig 5 ). It does not stain with carmine (compare the seminal vesicles with the organ in Fig. 5A View Fig 5 ), and only a few filaments are stained with hematoxylin ( Fig. 5B View Fig 5 ), so the presence of sperm cells is ruled out. We cannot entirely exclude the possibility of the presence of muscle cells because of a weak affinity with the fast green stain and because the few fibres that stain with hematoxylin have a strong affinity to the dye, comparable to that of the adjacent circular muscle fibres. However, this seems unlikely because the fibres run in random directions. Interestingly, at this level the longitudinal musculature of the body wall is interrupted, and only a few circular muscle fibres are present. No parenchymal gland cells were seen associated with this structure, either.
This fibrous mass was present in all the specimens, and its position, general organisation, and staining properties were constant; it is, therefore, unlikely to be an artefact. Given the absence of an analogous structure in the relevant bibliography, and for lack of a better term, we call it the 'fibrous organ'.
Taxonomic remarks
The following combination of morphological characters places the new species unequivocally in the genus Coelogynopora sensu Karling (1966) and Faubel & Rohde (1998): epidermis entirely ciliated, with intracellular nuclei; sclerotised copulatory system with stylet and spines (note that hard parts are variable among representatives of the genus, to the point that they may be absent); paired seminal vesicles located posteriorly to the gonopore; paired ovaries situated anteriorly to the pharynx, with two ovovitelline ducts that open separately into the posterior part of the genital atrium. While a bursa could not be discerned, a conspicuous female genital canal opens into the intestine through a genito-intestinal connection.
There are currently 39 known species of Coelogynopora ( Armonies 2018; Jouk et al. 2019), but, besides C. kenichii sp. nov., only three other species have all the spines fused proximally with the base of the penial stylet, namely Coelogynopora alata Tajika, 1981 , Coelogynopora brachystyla Karling, 1966 and Coelogynopora coniuncta Tajika, 1978 . Coelogynopora kenichii sp. nov. differs from all of them by having a different number of spines and by the shape of the penial stylet. Tajika (1981) reported 36 spines in C. alata ; in addition, the penial stylet has a more conical shape. Coelogynopora brachystyla has a single pair of spines; in addition, the penial stylet has a truncated end and bears a longitudinal slit that has been described as a “channel” ( Karling 1966). The closest species to C. kenichii sp. nov. is C. coniuncta . However, the penial stylet in the latter is longer than in C. kenichii sp. nov., and its base is much broader (the proportion of basal length to total length is 66% in C. coniuncta and 29% in C. kenichii sp. nov.); in addition, in C. coniuncta there are only four spines ( Tajika 1978, 1981). Besides the differences in the sclerotised copulatory system, in C. coniuncta the vasa deferentia enter the seminal vesicles through their anterior, ventral sides and not posteriorly as in C. kenichii sp. nov.; furthermore, in C. coniuncta the seminal vesicles extend posteriorly and, in most specimens, they were found to be fused at the level of the posterior end of the animal ( Tajika 1978).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |