
Chone infundibuliformis Krøyer, 1856
|
publication ID |
https://doi.org/10.5281/zenodo.171592 |
|
DOI |
https://doi.org/10.5281/zenodo.3508564 |
|
persistent identifier |
https://treatment.plazi.org/id/37244754-FF9B-FFB5-4A0E-FE6FFB7BFE84 |
|
treatment provided by |
Plazi |
|
scientific name |
Chone infundibuliformis Krøyer, 1856 |
| status |
|
Chone infundibuliformis Krøyer, 1856 View in CoL
Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , 4 View FIGURE 4 A–C, 5–11
Chone infundibuliformis Krøyer, 1856 View in CoL , 33.— Malmgren, 1866: 404 —405, Pl. 28, Fig. 87.— Malmgren, 1867: 116.— Cunningham and Ramage, 1887: 670, Pl. 44, Fig. 32.— McIntosh, 1916: 35, Pl. 2, Fig. 9 View FIGURE 9 .— Fauvel, 1927: 334, Fig. 116a–o.— WesenbergLund, 1950, 58.— Banse, 1972: 461 –465, Fig. 1 View FIGURE 1 a–l.— HartmannSchröder, 1996: 550, Fig. 168a–h.
Type material: Lectotype [ ZMUC POL –1749] Greenland, locality unknown, Leg. Krøyer. Paralectotypes [ USNM 376] Greenland, locality unknown, Leg. Krøyer, not Lutken (2). Paralectotypes [ BMNH 82.5.12.33] Greenland, locality unknown, Leg. Krøyer, not Lutken (3).
Nontype material: Greenland [ BMNH] 1921.5.1.4364, 4368/69 Greenland, Godhaven Harbour, “Valorous”, 9–18 m (5). 1921.5.1.4364, 4368/69 Greenland, Bressay Sound, Coll. Mc Intosh, 1921, 5, 1–436K (2). No number, Press. J. Berryman, Coll. Yell, Shetland, Whaal Firth in silt, 1979/80 (3). [ LACM – AHF] 0 0 3224 Goodhvan Fjord, dredged, Kristineberg Zool. Station (4). [ MCZ] North Atlantic Ocean, R/V Albatross IV, Cruise 9809, Sta. 187, July 29, 1998, NMFS 1999 (1). [ SMNH] 6861, 73073 W Greenland, Egedesminde, Leg. O. Torell (1). 6862, 73074 W Greenland, Godhavn, 55 m, 69° 14’ N, Leg. C. T. Amondsen (2). [ ZMB] 3106 Grönland, Karajak, Fjord. Leg. Vanhöffem S, July, 1893 (4); July 21, 1893 (1). 5167 Grönland, Coll. Grube/Oersted S. (3). 5168 Spitzbergen, Coll. Grube/Malmgren S. (1). [ ZMUC] POL –1761 Lille Hellefiskebanke, 4 miles W. S. W. of Rifkol, July 10, 1912, 40 m, Coll. Bornemann (1). POL –1764 East of Greenland, 4th Thule Expedition Sta. 26, Kangerolussuaq, August 20, 1933, 1–5 m (2). POL –1765 East of Greenland, 4th Thule Expedition Sta. 29, Kangerolussuaq, August 25, 1933, 4–5 m (28). Norway [ SMNH] 6864, 73076 Svalbard, Vest Spitzbergen, Hornsund, 73 m, Leg. Swedish Arctic Expedition, 1864 (2). [ ZMB] 1986 Spitzbergen, Kükenthal S. G. (2). [ ZMB] 6226 Spitzbergen, Exp. Rómer & Schaudinn S. Sta. 12 (1); Sta. 13 (3); Sta 14 (1); Sta. 36 (3); Sta. 41 (2); Sta 513 (2). Canada [ CMN] 3867 Byam Martin Channel, Sta. 10, 75 ° 01.4’ N, 106° 35’ W, R. K. S. Lee, July 14, 1974 (2). 3870 Byam Martin Channel, 74° 58.8’ N, 106° 23.5’ W, Sta. 2, Coll. R. K. S. Lee, June 18, 1974 (1). 1980–0171 New Foundland, Labrador Nain, 56° 36’ N, 62° N, Sta. SAR3–528B, Coll. J. Barrie, September 11, 1977 (1). 1982–0938 North West Territories, Davis Strait, 61° 10’ N, 61° W, Coll. Mac Laren Atlantic, October 23, 1976, Sta. 25 (3). 1988–0364 New Foundland, Labrador Shelf, Sta. 4, 53 ° 19’ N, 54° 41’ W, 350 m, Coll. D. Peer, Habitat Ecol Lab, September 5–6, 1987 (1). 1993–0030 Sta. B–2, Coll. Arctic Biol. Sta. Ste. Anne de Bellevue, 1949–00–00 (1). 19930031 Sta. 67–5. Coll. Arctic Biol. Sta. Ste. Anne de Bellevue, 1967–00–00 (3). 1993–0032 Frobisher Bay, Baffin, Sta. 5b, 63° 43.5’ N, 68° 31.7’ W, 13–23 m, Coll. Arctic Biol. Sta. Ste. Anne de Bellevue (2). 1995–0570 Sta. Theron, Th–451–45, Coll. Arctic Biol. Sta. Ste. Anne de Bellevue, September 0 1, 1965 (3). 1995–0571 Sta. Theron, Th–490 VVG–49, Coll. Arctic Biol. Sta. Ste. Anne de Bellevue, September 8, 1965 (5). NFLD Sta. 517, 51 ° 22’ N, 50° 21’31’’ W. Coll. S. W. Gorham, May 28, 1963, 360 m, Set. 134 (2). “Calanus” 1949 B–11 (1).
Additional material: Chone americana Day, 1973 [ FSBC EJ71414] Eastern Florida, St. Lucie County, Hutchinson Island, East of F. P. and L. Electrical generating plant, 27° 21’ 23’’ N, 80° 13’ 24’’ W, 11.2 m. Chone aurantiaca ( Johnson, 1901) , holotype [ MCZ 1933] Port Orchard, Puget Sound, Washington, Coll. A. Robertson, July 4, 1898. [ LACM – AHF] West Seattle, Washington, Vashon Island, Ferry dock, May 7, 2004, intertidal, Coll. B. Pernet. Chone farringtonae TovarHernández, 2005 , paratype [ FSBC I 66734] West Coast of Florida, off shore Pinellas County, Boca Ciega Bay, June 17, 1976. Chone georgesi Fitzhugh in press [ECOSUR] Isla Mujeres, Bajo Pepito, Mexico, in Stypopodium zonata, Coll. M. Díaz , February 1997. Chone gracilis Moore, 1906 , holotype [ USNM 5513] Alitak Bay, Kodiak Island, Alaska, Sta. 4274 (1903), 7.2– 42 m. Chone magna ( Moore, 1923) , holotype [ USNM 17281] Off Harris, Pt. San Miguel, California, Albatross R/V, Sta. 4436, April 15, 1904. Chone mollis ( Bush, 1904) , holotype [ YPM 2793] Harriman Alaska Expedition, Pacific Grove, California, Coll. W. R. Coe, 1901. Chone picta Verrill, 1885 , topotype [ YPM 30002] Martha’s Vineyard, Massachusetts. 70° 51’ N, 41° 22’ W. Coll. J. L. Simon and P. E. Schwamb, June 22, 1966. Chone princei McIntosh, 1916 , topotype [ CMN 1989–0394] Quebec, St. Lawrence estuary, off Point Mitis, 37 m, Sta. B610–7 IL –152D, Coll. Rafat MASSAD, July 0 9, 1971.
Redescription
The following redescription is based mainly upon the lectotype. Numbers in parentheses correspond to variation among paralectotypes and additional material.
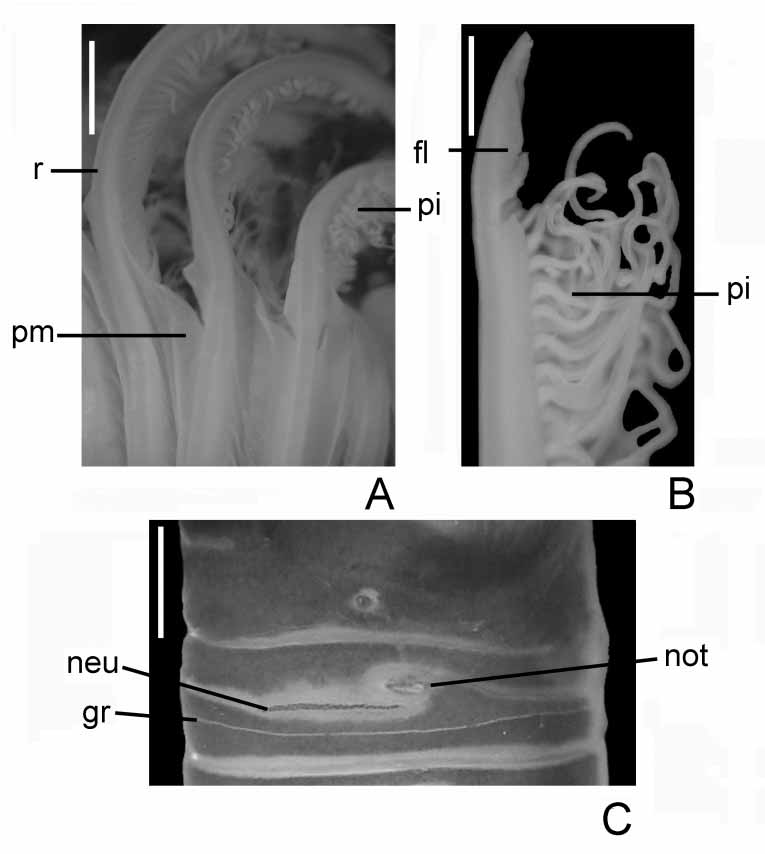
Colour, body shape and size: Body cream colored (specimen from North Atlantic Ocean pink when collected, MCZ, R/V Albatross IV), trunk cylindrical ( Fig. 1 View FIGURE 1 ). Body length: 81 mm (12–103), width: 8 mm (1–10). Tubes composed of fine sand.
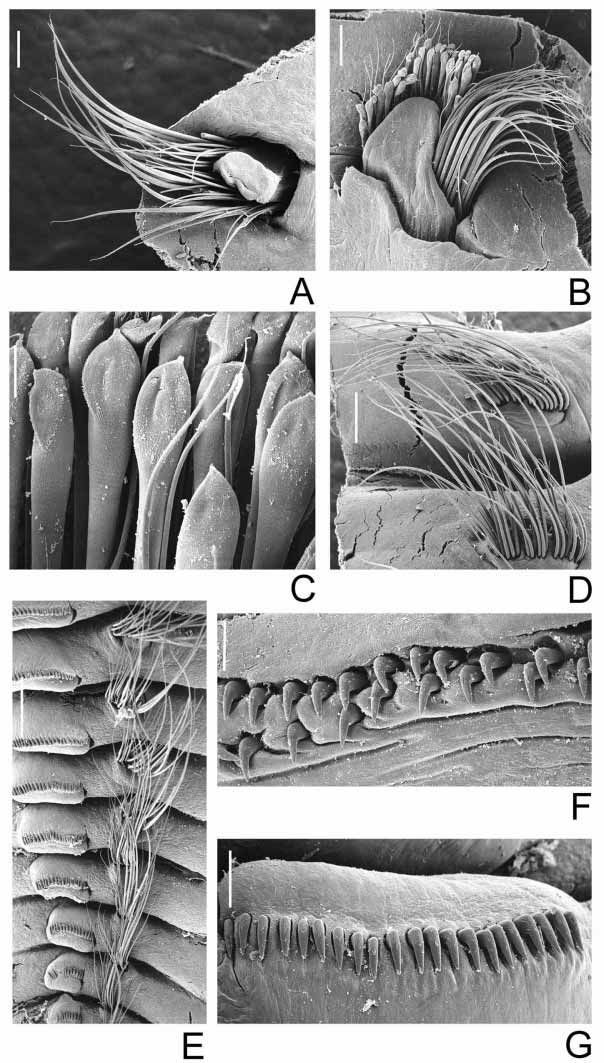
Branchial lobes and branchial crown: Branchial lobes mediumsized, located in lateral position on either side of mouth, curved such that they are concave in cross section relative to the mouth, insertion not exposed beyond collar ( Figs 1 View FIGURE 1 A–B). Branchial crown length: 19 mm (11–90). Radioles: 24 (7–23) pairs. Each radiole with paired series of pinnules of different size; median pinnules 3 times longer than more proximal pinnules. Radiolar tips: shortsized ( Fig. 2 View FIGURE 2 B). The palmate membrane extends up three quarters of the branchial crown length. Lateral flanges broad ( Figs 2 View FIGURE 2 A–B), outer radiole surfaces flat ( Fig. 2 View FIGURE 2 A). Dorsal lips extend from the inner, dorsal margins of the branchial lobes and terminate just dorsal to mouth ( Fig. 5 View FIGURE 5 A), broadly rounded in frontal view, as long as wide, without midrib, resembling the ventral lips ( Fig. 1 View FIGURE 1 I), longer than wide in dorsolateral view ( Figs 1 View FIGURE 1 E–F, H); dorsal lips vascularized by a plexus of small blood vessels derived from basal branchial blood vessels ( Fig. 6 View FIGURE 6 A–B); filled with hyaline cartilage ( Figs 7 View FIGURE 7 A–C, E), no extension of branchial skeleton. Dorsal margin of dorsal lips extends up along inner margin of dorsalmost radiole, fused with 8 ( 5–9–10) dorsal pinnular appendages united by a palmate membrane ( Fig. 1 View FIGURE 1 E). Ventral lips rounded, as long as wide, about half of the dorsal lips length, extending from the central margin of the branchial lobes ( Fig. 1 View FIGURE 1 H), filled with hyaline cartilage ( Figs 8 View FIGURE 8 A–C). Ventral radiolar appendages: 6 (3–7) pairs of similar length, about one quarter of the branchial crown length, originate just ventral to ventral lips ( Figs 9 View FIGURE 9 A–B), continuous with ventral extensions of inner margins of branchial lobes ( Fig. 1 View FIGURE 1 H).
Peristomium: Anterior peristomial ring lobe not exposed beyond collar, distally entire, triangular. Posterior peristomial ring collar: anterodorsal, lateral and ventral margins entire, ventral margin slightly higher than dorsal ( Figs 1 View FIGURE 1 A–B), entire length of middorsal collar margins form a narrow gap. Ventral shield of collar swollen, horseshoeshaped, two times wider than long; ventral shield divided transversally by a white and slender line, the anterior half transparent ( Fig. 1 View FIGURE 1 A). The posterior peristomial ring collar length is as long as 1.5 times the length of chaetiger 2 in lateral view.
Thorax: Chaetiger 1: Notopodia: two groups of 14–16 chaetae, chaetae from the exterior group twice longer than interior group ( Fig. 3 View FIGURE 3 A). Chaetigers 2 to 8: Notopodia: superior group with two irregular rows of elongate, narrowly hooded chaetae, each row with 30 (26–42) chaetae ( Fig. 3 View FIGURE 3 B); inferior group with one anterior row with 16 (18–28) short bayonet chaetae ( Figs 3 View FIGURE 3 B–C), 2 posterior rows with 22 (31–43) symmetrical, paleate chaetae with short mucro ( Figs 3 View FIGURE 3 B–C). Pre and postchaetal lobes well developed ( Fig. 3 View FIGURE 3 B). Neuropodia: 29 (39–45) acicular uncini per tori, distributed as an irregular double row (except in juveniles); heads in the same direction ( Fig. 3 View FIGURE 3 F); the oldest upper parts of the tori have only one row (one quarter of the tori length); main fang surmounted by four rows of teeth in frontal view, occupying less than half of the main fang length; teeth unequal in size ( Fig. 4 View FIGURE 4 A). Biannulate condition in thoracic segments is given by the presence of well differentiate intersegmental grooves, and internotopodial and interneuropodial grooves, less differentiated than intersegmental ( Figs 1 View FIGURE 1 A–B). Glandular ridge on chaetiger 2, narrow, just below chaetal lobe and tori ( Figs 1 View FIGURE 1 A–B, 2C, 11A, F–G).
Abdomen: Abdominal segments: 44 (28–76). Anterior segments: two transverse rows of 22 elongate, narrowly hooded chaetae, chaetae from the upper row half as long as chaetae in lower row ( Figs 3 View FIGURE 3 D–E); 25 (29–33) uncini per tori ( Fig. 3 View FIGURE 3 G), older and younger uncini with the main fang surmounted by five rows of teeth in frontal view, occupying less than half of the main fang length ( Fig. 4 View FIGURE 4 B), main fang not extending beyond breast ( Fig. 1 View FIGURE 1 G). Posterior segments: 23–28 very elongate, narrowly hooded chaetae, 25% longer than those of anterior segments ( Fig. 3 View FIGURE 3 E); 3 (3–4) uncini per torus, similar to anterior segments, but shorter ( Figs 1 View FIGURE 1 J, 4C). Intersegmental grooves well differentiate, internotopodial and interneuropodial grooves less differentiated than intersegmental. Pygidium with rounded posterior margin ( Fig. 1 View FIGURE 1 D).
Gametes: Lectotype is a female, oocytes in all thoracic and abdominal segments, diameter 6.07 m. Specimens from CMN 3870 are females with oocytes distributed in all thoracic and abdominal segments; males with sperm in anterior abdominal chaetigers, spermatids develop in tetrads, and mushroomshaped acrosom.
Methyl green staining: The epidermis is completely glandularized and stains uniformly in thorax and abdomen, dorsal and ventrally, except in except intersegmental grooves, internotochaetal and neurochaetal grooves, and both pre and postchaetal lobes ( Figs 1 View FIGURE 1 A–B). The collar segment is darker in the anterior half, except for anterior end of the ventral shield of collar.
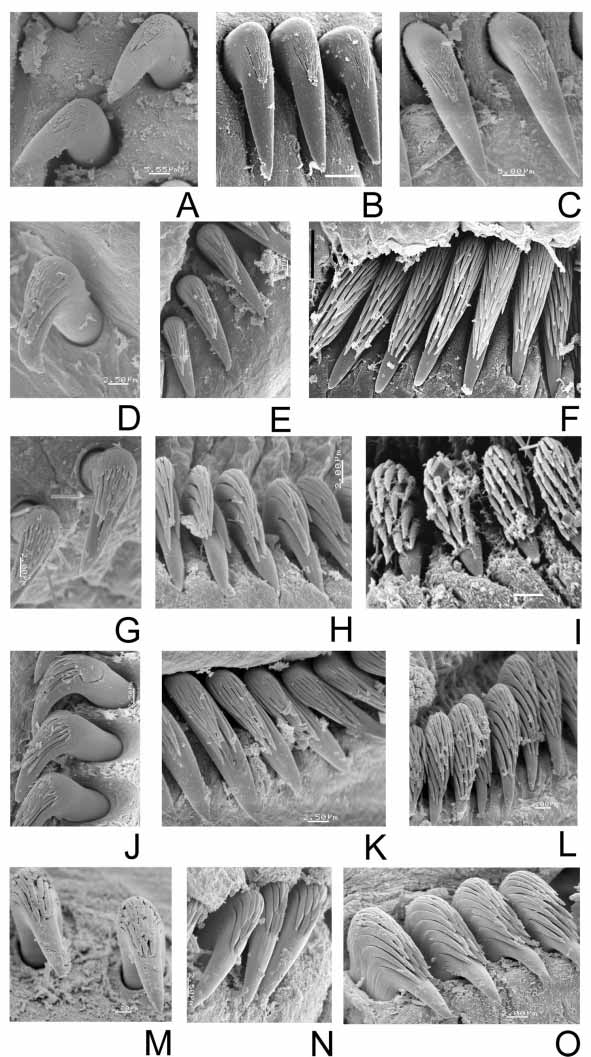
Remarks: Scanning electron microscopy revealed an important character in this species: teeth above the main fang of thoracic acicular uncini are of unequal size ( Fig. 4 View FIGURE 4 A). Fitzhugh (1989) thought that all the teeth above main fang were of the same size, and that the presence of unequalsized teeth was a synapomorphy for Amphicorina Claparède, 1864 . This character is, however, homoplastic, occurring in most of the fabriciin genera, and its presence is now documented in C. infundibuliformis , C. georgesi ( Fig. 4 View FIGURE 4 D), C. americana ( Fig. 4 View FIGURE 4 G) and C. farringtonae ( Fig. 4 View FIGURE 4 J). In C. infundibuliformis , the only feature that tends to be dependent on size and age is the presence of thoracic uncini distributed as irregular double row in adults ( Fig. 3 View FIGURE 3 F), while in juveniles there is only one row of uncini per torus.
The presence of modified uncini in posterior abdominal segments as raspshaped plates or Amphicorina type (see Rouse 1994: Fig. 17) have been recorded or illustrated in some species of Chone : C. longiseta Giangrande, 1992 , C. acustica Claparède, 1870 , and C. filicaudata Southern, 1914 (in Giangrande 1992); C. trilobata Gallardo, 1968 , C. sp.1, C. sp. 2, C. sp. 3, C. sp. 4 and C. sp. 6 (in Fitzhugh, 2002); C. americana Day, 1973 , C. diazi , C. farringtonae , C. johnstonae , C. perkinsi and C. uebelackerae ( TovarHernández, 2005) . This kind of modified uncini ( Figs 4 View FIGURE 4 F, I, L, O) have a main fang surmounted by several regular, vertical rows of small teeth of equal size that occupy at least three quarters of the main fang length, and a poorly developed rectangular or subrectangular breast, instead of a few rows of unequalsized teeth occupying less than half of the main fang length and a well developed breast, as anterior segment uncini ( Fig 4 View FIGURE 4 E, H, K, N). However, anterior and posterior abdominal uncini are similar in shape in the type species. They possess a well developed main fang surmounted by a few smaller teeth that occupy less than half of its length ( Figs 4 View FIGURE 4 B–C) and are irregularly arranged in an overlapping pattern, instead of a uniform set of rows ( Fig. 4 View FIGURE 4 C). The breast is well developed and main fang does not extend beyond it. Other species reviewed with anterior and posterior abdominal uncini of similar shape include: C. aurantiaca [MCZ 1933, holotype], C. gracilis [USNM 5513, holotype], C. magna [USNM 17281, holotype], C. mollis [YPM 2793, holotype] and C. picta [YPM 30002].
In those species of Chone with modified, posterior abdominal uncini, the presence of a posterior, lateroventral depression is easily seen (see TovarHernández 2005, Fig. 11 View FIGURE 11 B); however, this depression is not the same as the anal depression in Euchone Malmgren, 1866 . In Euchone , the anal depression (=anal plate, funnel, spoonshaped cavity or scoop) is ventral and extends from the pygidium through a varying number of abdominal chaetigers ( Fitzhugh 1989). Cochrane (2003) recognized the anal depression as either being bordered by lateral wings or clearly demarcated by anterior and/or lateral ridges; she also detected a degree of ambiguity between a posteriorly enlarged faecal groove or dorsoventral depression in certain Chone and Amphicorina species, and a poorly developed anal depression in some Euchone . In species of Euchone , the Amphicorina type abdominal uncini are primarily located in the area of the anal depression ( Banse 1970; Fitzhugh 1989); in species of Chone with modified abdominal uncini ( C. georgesi ( Fig. 4 View FIGURE 4 F), C. americana ( Fig. 4 View FIGURE 4 I), C. farringtonae ( Fig. 4 View FIGURE 4 L) and C. princei ( Fig. 4 View FIGURE 4 O)), they are also located in the segments of the lateroventral depression.
Histology
Gross observations
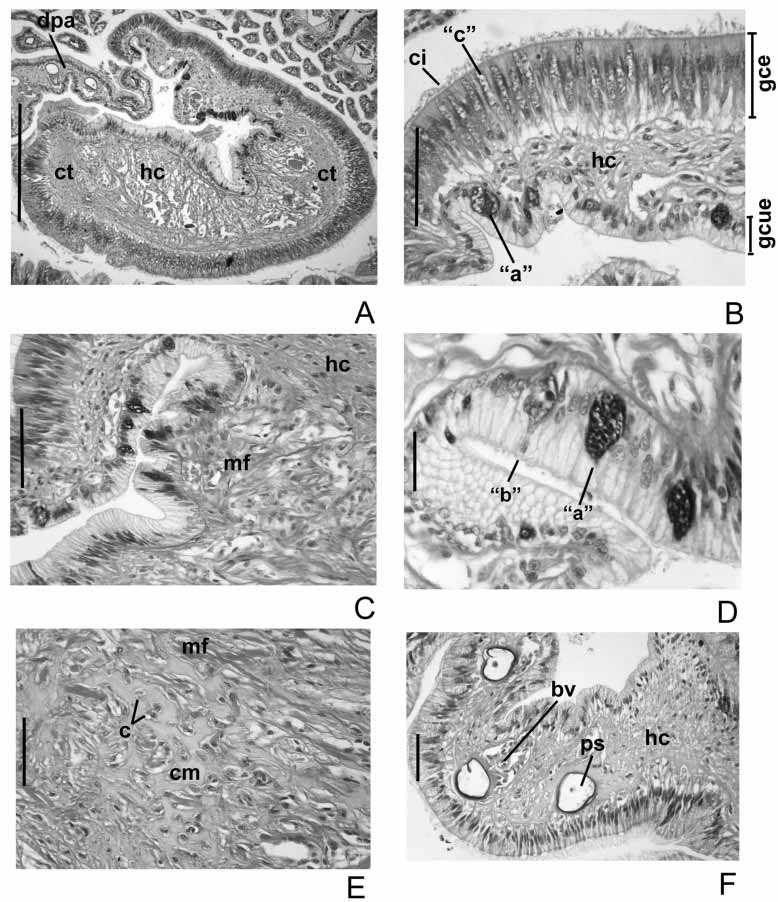
The branchial crown in C. infundibuliformis is supported basally by a central mass of cartilaginous skeleton and two lateral bars or horns of cartilaginous skeleton, both completely separated, independent one of another. i) The basal central mass of cartilaginous skeleton ( bcs) is located above the subesophageal ganglia ( Figs 5 View FIGURE 5 A, 6A), divided basally and covered anterolaterally by smooth muscle ( m) ( Fig. 6 View FIGURE 6 A). ii) The lateral bars or horns of cartilaginous skeleton ( bls) are composed of a large mass of cells in the base of the branchial lobes extending through radioles as two rows of cells ( Figs 5 View FIGURE 5 A, 6A).
The branchial crown is vascularized by two large blood vessels ( bv) ( Figs 5 View FIGURE 5 A, 6A); each vessel is covered by a coelomic chamber ( cc) and conjunctive tissue ( ct) ( Fig. 5 View FIGURE 5 C). The branchial crown is innervated ( n) at the base of each branchial lobe ( Figs 5 View FIGURE 5 A, D, 6A) and supported with hyaline cartilage on lateral sides of mouth ( m) ( Fig. 5 View FIGURE 5 A). The hyaline cartilage is constituted of chondrocytes ( c) and a homogeneous eosinophilic matrix ( cm) ( Fig. 5 View FIGURE 5 E). Some muscular fibers ( mf) are disposed in dense bundles in different directions ( Fig. 5 View FIGURE 5 B).
Dorsal lips and dorsal pinnular appendages
The dorsal lips are vascularized by a plexus of small blood vessels ( bv) than run along the lip as strongly branched network ( Figs 6 View FIGURE 6 B–C, 7A); each blood vessel is surrounded by coelomic chambers ( cc) ( Figs 6 View FIGURE 6 B–C). These blood vessels are derived from the basal branchial blood vessels, which further back communicate with the dorsal vessel ( Fig. 6 View FIGURE 6 A). The dorsal lips have hyaline cartilage, but there is no extension of the branchial skeleton. The hyaline cartilage ( hc) extends into the middle region of the dorsal lip and along the inner lip margin ( Fig. 7 View FIGURE 7 A); it is composed of abundant collagen fibers ( cf) forming a net, and spread condrocytes ( c) ( Figs 7 View FIGURE 7 A–C, E). The dorsal lips have conjuntive tissue ( ct) extending along the outer margin of the lip ( Figs 6 View FIGURE 6 B–C, 7A). Two types of simple epithelium are present in dorsal lips: columnar ( gce) and cubic ( gcue) ( Fig. 7 View FIGURE 7 B). The columnar epithelium covers the external/outer margin of the lips ( Figs 6 View FIGURE 6 B–C, 7B), includes ciliated cells ( ci) ( Figs 6 View FIGURE 6 B–C) and type “c” glands ( Fig. 7 View FIGURE 7 B), and is covered by cuticle ( cu) ( Figs. 6 View FIGURE 6 B–C, 7B). The cubic epithelium covers the internal/inner margin of the lip ( Fig. 7 View FIGURE 7 B), there are not ciliated cells ( Fig. 6 View FIGURE 6 B–C), but contains types “a” and “b” glands, and is covered by cuticle ( Figs 7 View FIGURE 7 B, D).
The external pinnula of each dorsalmost radiole is fused with six pinnules ( dpa) towards the dorsal, central margin of the branchial crown, above of the basal, central skeleton ( Figs 5 View FIGURE 5 A, 6A); they are united by a glandular, columnar epithelium composed of ciliated and types “b”, “c”, “d” glands. Each dorsal pinnular appendage contains a central blood vessel surrounded by the coelom, and associated with the pinnular skeleton and hyaline cartilage ( Fig. 7 View FIGURE 7 F).
Ventral lips and ventral radiolar appendages
The ventral margins of the ventral lips are each fused with three undeveloped radioles ( unr) ( Fig. 8 View FIGURE 8 A–B). The ventral lips are composed of hyaline cartilage ( hc) ( Figs 8 View FIGURE 8 A–C) and ciliated epithelium and type “c” glands.
Radioles
The epithelium of radioles is simple columnar, covered by cuticle ( cu) and composed of type “c” glands ( Figs 9 View FIGURE 9 A–B). One pair of nerves ( n) is between each pair of radioles ( Fig. 9 View FIGURE 9 A). In cross section, the radiolar skeleton ( rs) is covered by hyaline cartilage, composed of a dense cartilage matrix ( cm) with few chondrocytes ( c) ( Fig. 9 View FIGURE 9 C); in transversal section, the radiolar skeleton ( rs) continues into each pinnule ( ps), which is divided by longitudinal muscle ( m) and conjunctive tissue ( ct) ( Fig. 9 View FIGURE 9 D). Directly from the branchial blood vessel, which further back communicates with the dorsal vessel, a small blood vessel branches off to each radiole and then, into each pinnule, surrounded by a coelomic chamber ( cc) ( Fig. 9 View FIGURE 9 E). In cross section, each radiole is composed of a central endoskeleton axis consisting of two large cartilaginous cells ( rs) ( Fig. 9 View FIGURE 9 A), longitudinal muscle ( m) and two pairs of nerves ( n) running along the basal groove, and ciliated epithelium in the basal groove of the radiole ( ci) ( Figs 9 View FIGURE 9 A, D–E).
Collar
The midventral margin of the posterior peristomial ring collar consist of a broad central axis of hyaline cartilage ( hc), including the area of the ventral shield of collar ( vsc) ( Fig. 10 View FIGURE 10 A); composed of a dense cartilage matrix ( cm) with few chondrocytes ( Figs 10 View FIGURE 10 A, D–E) and covered on both sides by glandular epithelium with types “a”, “b” and “c” glands (markedly basophilic and substantially larger) and cuticle ( cu) ( Figs 10 View FIGURE 10 A–C).
Epithelium and glandular ridge on chaetiger 2
The epithelium of Chone infundibuliformis has type “a”, “b” and “c” glands markedly basophilic and substantially larger, covered by cuticle ( cu) ( Figs11 View FIGURE 11 A, D–F). Each intersegmental division is reinforced with conjunctive tissue ( ct) and nerves ( n) ( Fig. 11 View FIGURE 11 B). At the base of the epithelium, there are several nerves ( n) and blood vessels ( bv) ( Figs 11 View FIGURE 11 A, C). The glandular ridge in C. infundibuliformis and C. aurantiaca occupies only the external half of the epithelium; it is composed of strongly differentiated acidophil glandular cells ( ag), tubularshaped with granulose secretions ( Fig. 11 View FIGURE 11 A, G, 12A–D).







| ZMUC |
Zoological Museum, University of Copenhagen |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| LACM |
Natural History Museum of Los Angeles County |
| AHF |
Allan Hancock Foundation, University of Southern California |
| MCZ |
Museum of Comparative Zoology |
| SMNH |
Saskatchewan Museum of Natural History |
| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
| CMN |
Canadian Museum of Nature |
| YPM |
Peabody Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chone infundibuliformis Krøyer, 1856
| Tovar-Hernández, María Ana & Sosa-Rodríguez, Teresa 2006 |
Chone infundibuliformis Krøyer, 1856
| Hartmann-Schroder 1996: 550 |
| Banse 1972: 461 |
| Fauvel 1927: 334 |
| McIntosh 1916: 35 |
| Cunningham 1887: 670 |
| Malmgren 1867: 116 |
| Malmgren 1866: 404 |