
Chelonoidis marcanoi, Chapman, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4277.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:FEA3BBA6-1BAA-4584-932F-B2FE5BC2C648 |
|
DOI |
https://doi.org/10.5281/zenodo.6051677 |
|
persistent identifier |
https://treatment.plazi.org/id/03A4E650-0B0A-FFEC-FF5A-F8482DD4A35D |
|
treatment provided by |
Plazi |
|
scientific name |
Chelonoidis marcanoi |
| status |
sp. nov. |
Chelonoidis marcanoi sp. nov.
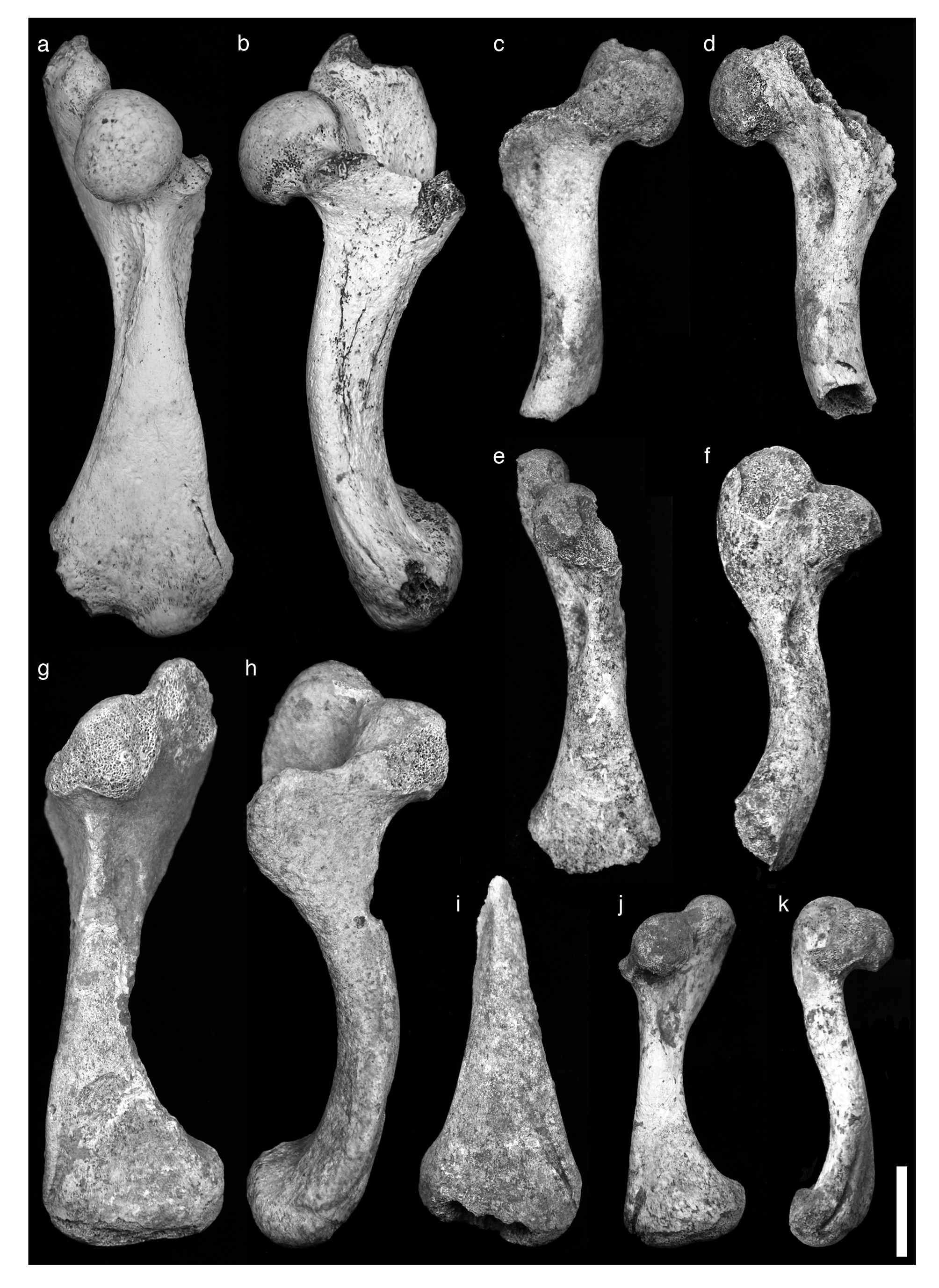
Holotype. Right humerus (NHMUK PV R 36954), collected on 12 May 2007 ( Figure 2a–b View FIGURE 2 ).
Paratypes. NHMUK PV R 36955; MNHNSD FOS 23.1054-23.1064; UF 26095-26100.
Type locality. Cueva del Papayo, Pedernales Province, Dominican Republic ( NHMUK PV R 36954-36955).
Other known localities. Unnamed cave near Bayaguana, Monte Plata Province , Dominican Republic (UF 26095-26100); Cueva No . 12 ( MNHNSD FOS 23.1054-23.1056 ), Cueva de Las Tortugas ( MNHNSD FOS 23.1057-23.1062 ), Cueva del Muerto ( MNHNSD FOS 23.1063 ) and Cueva de las Caritas ( MNHNSD FOS 23.1064 ), Pedernales Province , Dominican Republic. Only known from Hispaniola.
Etymology. After Professor Eugenio de Jesús Marcano, who collected the first giant tortoise fossils discovered on Hispaniola that were described by Franz & Woods (1983).
Diagnosis. Thin-shelled, large-bodied testudinid tortoise with adult humerus much larger than in C. alburyorum but probably smaller than in C. sombrerensis ; humerus more slender than in C. alburyorum or C. cubensis ; M. latissimus dorsi muscle scar on lateral surface better defined than in C. monensis but not as prominent as in C. cubensis ; humeral shaft rounded/oval or slightly triangular and approximately as long as wide in middle shaft cross-section, unlike the narrower, deeper shaft of C. cubensis . Femur with maximum distal width markedly greater than maximum proximal width in single available young specimen, versus the equal proximal and distal widths shown by C. alburyorum . External carapace surface showing sulci with edges raised as distinct low ridges, unlike the indistinct and scarcely apparent sulci of C. monensis , C. sombrerensis , and the undescribed giant tortoises from Navassa and New Providence; gular scutes much shorter than in C. cubensis , with sulci defining their posterior border curving posteriorly rather than anteriorly as in C. alburyorum .
Description. Humerus: Shaft straight along proximal half, recurved ventrally along distal half; relatively long and slender. Proximal width across trochanters and distal width across articulating surface approximately equal. Head subspherical, slightly laterally compressed, oriented anteriorly at almost 90°. Trochanteric tuberosities straight, diverging proximally at about 40°; posterior edges raised up as narrow laterally flattened ridges at their base, becoming more rounded towards proximal surfaces. Intertrochanteric fossa short, deep, with roughened surface at distal end. Lesser trochanter short; proximal surface flattened, almost level with midline of humeral head in well-preserved specimens. Greater trochanter much longer, extending considerably beyond upper surface of humeral head; proximal surface rounded. Prominent, relatively deep and oval M. latissimus dorsi muscle scar welldefined on lateral surface of humerus in front of base of greater trochanter; with roughened inner surface. Second much shallower circular muscle scar also present on medial surface of humerus, more anteriorly situated than M. latissimus dorsi muscle scar. Sides of shaft flattened immediately distal to humeral head, with anterior surface raised as narrow ridge between medial and lateral muscle scars; anterior surface becoming broader distally to form smooth, posteriorly curved and mediolaterally flattened surface across distal half of humerus. Medial surface approximately straight; lateral surface curving outwards distally, giving flared outline to humerus in anterior profile. Shaft midlength rounded/oval or slightly triangular in cross-section, approximately as long as wide. Distal anterior surface with prominent long, straight, deep and narrow incised entepicondylar groove close to medial margin, running mediodistally up to or into the medial corner of the articular surface. Posterior distal surface with two short, low raised ridges, which diverge towards distal articulating surface. Distal articulating surface of holotype developed into two low, swollen facets that correspond to the ulnar and radial articulating surfaces; facets less developed in other specimens. Humeral measurements given in Table 1.
Femur: Proximal morphology only available for single small specimen of young individual with spongy, incompletely formed articular surfaces (MNHNSD FOS 23.1063); adult distal femoral morphology displayed by second, incomplete specimen (MNHNSD FOS 23.1055). Femoral head damaged; probably originally spherical. Trochanters low, not extending above level of femoral head; greater trochanter slightly higher than lesser trochanter, with more rounded proximal surface. Trochanters enclose deep, rounded medial trochanteric fossa on proximal surface. Shaft flat in medial/lateral profile, with distal articulating surface curving posteriorly along distal quarter of shaft. Prominent ovoid muscle scar present on lateral shaft surface below femoral head, about one-third length along shaft. Shaft midpoint robust and approximately square in cross-section. In dorsal profile, shaft narrows slightly below head then flares laterally and expands in width across distal half of shaft, so that maximum distal width is markedly greater than maximum proximal width, and anterior surface of distal shaft is broad and flattened. No prominent grooves on distal anterior surface. Distal articulating surface shows very weak development of facets on adult specimen, defined by faint notch and corresponding to tibial and fibial articulating surfaces. Femoral measurements given in Table 2.
Plastron and carapace: Epiplastron represented by two fragments (MNHNSD FOS 23.1056, 23.1060) that are identifiable from their deeply scalloped natural border, thin non-triangular cross-section, and lack of insertion of costal plates or ribs. These fragments represent two individuals from different sites. External surface of epiplastron slightly rounded. Gular apices consist of two gently anteriorly rounded lobes, separated at midline by shallow notch. Sulci defining gular scutes on external surface expressed as simple shallow unraised grooves. Gular scutes short, separated at midline by sulcus measuring 17.9 mm in length in MNHNSD FOS 23.1056, and 18.6 mm in length in MNHNSD FOS 23.1060. Sulci defining posterior border of gular scutes first curve gently outwards posterolaterally from midline, then curve more posteriorly as they expand. Inner surface inflated towards skin line; surface behind skin line slopes sharply from epiplastral shelf. Shell very thin; skin line thickness measures 15.5 mm in MNHNSD FOS 23.1056, and 16.0 mm in MNHNSD FOS 23.1060; behind skin line, shell thickness narrows to c. 3 mm at inner margin of MNHNSD FOS 23.1056, and 4.2 mm at inner margin of MNHNSD FOS 23.1060. No external or internal ornamentation (e.g. “bird face” of Franz & Franz 2009) seen on either available epiplastron fragment. Peripheral fragment (MNHNSD FOS 23.1061) thicker and distinctly triangular in crosssection in comparison to epiplastral fragments, and with very gently lobed margin; skin line thickness measures 20.5 mm, narrowing to 3.8 mm at inner margin. Fragment comprising two successive costal plates (NHMUK PV R 36955) with curved external and internal surfaces; maximum thickness of 11.0 mm, minimum thickness of 4.5 mm. No external ornamentation; internal surface with two low raised parallel ridges associated with development of costal ribs. Fragment from border of carapacial rim (MNHNSD FOS 23.1062) consists of axillary buttress supporting right anterior corner of carapace-plastron bridge at a position immediately behind the buttress, and includes two peripherals and part of a costal plate; maximum buttress thickness 21.7 mm; lateral margin very gently scalloped; carapace with a minimum thickness of 2.9 mm, and with external surface showing sulci with edges raised as low ridges. Additional description of carapace fragments previously given by Franz & Woods (1983).
Remarks. We interpret our new collection of giant tortoise fossils as all representing a single species, as the multiple humeri in this collection exhibit no morphological differences other than ontogenetic variation, and the fossils were all preserved in comparable depositional conditions with similar co-occurring vertebrate faunas, suggestive of similarity in geological age. The humeri in our new collection are also indistinguishable from the single chelonian humerus from Bayaguana previously described and illustrated by Franz & Woods (1983), and our new carapace and plastron fragments show identical morphological characteristics to the Bayaguana material, so we refer all of the known giant tortoise specimens from Hispaniola to the same new species.
Although carapace and neural characteristics that can be used to differentiate Chelonoidis from other New World testudinid genera (cf. Franz & Franz 2009) cannot be determined from available material of the new species, several characters of the humerus (the best-preserved and most commonly represented element in our collection) support genus-level assignment of the new species to Chelonoidis rather than to North American xerobatin testudinids ( Gopherus , Hesperotestudo ), the other candidate genera to which the extinct Caribbean tortoises might be referable ( Williams 1952; Auffenberg 1974; Meylan & Sterrer 2000). Unlike the Hispaniolan giant tortoise, mainland Neotropical Chelonoidis species or C. alburyorum , the humeri of Gopherus and of mainland and insular Hesperotestudo species lack either an obvious insertion site for M. latissimus dorsi , or an entepicondylar groove or foramen; the greater trochanter also does not extend beyond the head of the humerus in Hesperotestudo , in contrast to the marked extension of the greater trochanter in the Hispaniolan giant tortoise and other Chelonoidis species, and this character is interpreted as a derived condition in Chelonoidis ( Auffenberg 1963; Meylan & Sterrer 2000; Franz & Franz 2009). The shell of the insular Hesperotestudo bermudae is also not markedly thinner than in mainland American xerobatin testudinids ( Meylan & Sterrer 2000), in contrast to the very thin-walled shell seen in the Hispaniolan giant tortoise and other described Chelonoidis species ( Franz & Franz 2009). Whilst variation in shell thickness may represent an ecological response to insular environmental conditions (e.g. absence of predatory mammals) rather than a phylogenetic signal, the shell state in the Hispaniolan tortoise is consistent with that described for species of Chelonoidis .
We therefore consider that even in the absence of information on skull morphology or detailed shell morphology, the Hispaniolan tortoise can be confidently referred to Chelonoidis on the basis of comparative postcranial characteristics. This taxonomic hypothesis is consistent with other available data on Quaternary testudinid biogeography for the insular Caribbean, as Chelonoidis definitely occurred on the nearby Bahamian Archipelago ( Kehlmaier et al. 2017). The only potentially unusual characteristic shown by the Hispaniolan tortoise compared to well-described Chelonoidis species is the apparent absence of diagnostic interior sculpture on entoplastral elements in the new collection, but this may simply reflect the limited preservation of available incomplete entoplastral specimens from the new species.
The Hispaniolan species differs from C. alburyorum , the best-described Caribbean giant tortoise, in being considerably larger in body size. Although the total shell length is unknown for the Hispaniolan species, Franz & Woods (1983) estimated an approximate total carapace length of c. 600 mm through comparison with modern giant tortoise specimens of known size, and available adult tortoise humeri from Hispaniola have a total length of c. 125– 135 mm ( Table 1; Franz & Woods 1983); in contrast, C. alburyorum has a total carapace length of 345–466 mm, and a humerus length of 96.1 mm ( Franz & Franz 2009). The humerus of the Hispaniolan species is also more slender and gracile than in C. alburyorum ; despite being markedly longer, the maximum width across the distal articulating surface in well-preserved adult Hispaniolan humeri is only 38.6–40.2 mm (30.7% of total humerus length in MNHNSD FOS 23.1054), compared to 37.4 mm in C. alburyorum (38.9% of total humerus length) ( Franz & Franz 2009). Femoral comparisons between the two taxa are based on more limited available material, but the single complete femur of the Hispaniolan species has a maximum distal width that is markedly greater than its maximum proximal width, in contrast to the similar proximal and distal widths seen in femora of C. alburyorum . The available Hispaniolan femur is from a young individual, however, so it is possible that this difference may at least partly reflect ontogenetic variation. The Hispaniolan species also apparently lacks the diagnostic interior “bird face” sculpture seen on the entoplastron of C. alburyorum , and has shorter gular scutes defined on the external surface by sulci which curve outwards and gently backwards rather than running outwards and forwards from the plastron midline as in C. alburyorum .
Detailed comparison with other Caribbean giant tortoises is more difficult due to the limited material available from other islands, but diagnostic differences can also be identified between the Hispaniolan species and other previously described Caribbean tortoise taxa. Humeral measurements have not been reported for C. cubensis or C. monensis , but C. cubensis differs from the Hispaniolan species in having a shorter, more robust humerus with a shaft depth c.1.5 times the shaft width and that shows a much more strongly developed, extremely large and deep M. latissimus dorsi muscle scar, and also in having an extremely elongate gular scute and a prominent “bird face” sculpture on the entoplastron ( Williams 1950; Franz & Franz 2009); and C. monensis conversely differs from the Hispaniolan species in reportedly having a reduced M. latissimus dorsi muscle scar that merely consists of a roughened area rather than a pit ( Williams 1952). Chelonoidis sombrerensis is much more poorly known; one described humerus is similar in length to that of the Hispaniolan species (estimated at approximately 130 mm), but this specimen lacks both ends and so may not be fully grown; another reported humerus for this species is markedly larger (approximately 300 mm), and with a considerably greater shaft depth than in the Hispaniolan species (1.3 times the shaft width) ( Julien 1878; Williams 1952). The Hispaniolan species also differs from C. monensis , C. sombrerensis , and the undescribed “moderate-sized” tortoises from Navassa and New Providence in having carapace sulci with raised ridges rather than having sulci that are indistinct and scarcely apparent ( Auffenberg 1967); carapace sulci defined by raised edges along their margins are otherwise only shown by C. alburyorum and C. cubensis in the Caribbean region ( Williams 1950; Franz & Franz 2009).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |