
Chaetacanthus brasiliensis ( de Quatrefages, 1866 ) Salazar-Silva & López-Sánchez & Salazar-Vallejo, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4885.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:AFE72E8B-A590-4B89-96A1-880C2D2AE14B |
|
DOI |
https://doi.org/10.5281/zenodo.4329440 |
|
persistent identifier |
https://treatment.plazi.org/id/03EC4719-FFCF-E551-66D0-FA30FBA864E7 |
|
treatment provided by |
Plazi |
|
scientific name |
Chaetacanthus brasiliensis ( de Quatrefages, 1866 ) |
| status |
comb. nov. |
Chaetacanthus brasiliensis ( de Quatrefages, 1866) n. comb.
Figures 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , 7 View FIGURE 7 , 8 View FIGURE 8 , 13 View FIGURE 13 D–F
Polynoe brasiliensis de Quatrefages, 1866: 246–247 View in CoL .
Iphione magnifica Grube, 1876: 51 View in CoL .
Polynoe branchiata Treadwell, 1901:186 View in CoL ; Horst 1922: 197–198, Fig. 1 View FIGURE 1 .
Lepidonotus (Physalidonotus) barbatus Augener 1910: 244–246 View in CoL , Figs 4–6 View FIGURE 4 View FIGURE 5 View FIGURE 6 .
Lepidonotus (Physalidonotus) magnificus: Augener 1922: 40 .
Lepidonotus brasiliensis: Seidler 1924: 37 View in CoL (n. comb.); Amaral & Nonato 1982: 25; de Assis et al. 2015: 65 View Cited Treatment (Tabl. 1), 68.
Physalidonotus magnificus: Seidler 1921: 87 (n. comb.).
Chaetacantus magnificus: Seidler 1922: 301 ; Seidler 1924: 97 (n. comb., syn.); Amaral & Nonato 1982: 18 –19, Figs 24–31; de Brito et al. 2013: 27–29, Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 .
Euphione (Chaetacanthus) magnificus: Augener 1927: 43–44 .
Lepidonotus branchiatus Treadwell, 1939: 183–184 View in CoL , Fig. 16 View FIGURE 16 .
Lepidonotus pilosus: Treadwell 1941: 26 View in CoL (specimen not seen; non Treadwell, 1937).
Type material. Brazil. Holotype of Polynoe brasiliensis de Quatrefages, 1866 , MNHN 78 About MNHN , Bahía, 1868 . Trinidad. Holotype of Iphione magnifica Grube, 1876 , ZMB 1059 View Materials , Schilling leg. (plus one slide made by Seidler). Puerto Rico. Holotype of Polynoe branchiata Treadwell, 1901 , USNM 16006 About USNM , Harbor Mayaguez , Bahía Boquerón, United States Fish Commission Steamer Fish Hawk, Sta. 6065, 8– 10 m, 20 Jan. 1899.
Additional material. Brazil. UMML 4215 View Materials , R/V Oregon, Cruise 84, Sta. 4215 (00°27’ N, 47º09’ W), 22 m, 26 Feb. 1963 [3.5 cm long, 1.2 cm wide, 26 segments]. GoogleMaps Caribbean Panama. Four specimens ( ECOSUR P3100 View Materials ), Bocas del Toro, Islote Manglar , off Isla Bastimento , 2 m, dead Agaricia coral , 2 m depth, 12 Jul. 2018, O. Molina, coll. [2.5–4.5 mm long, 0.7–1.5 mm wide (without chaetae), 26 segments; nephridial lobes from segment 7] GoogleMaps .
Diagnosis. Chaetacanthus with branchial filaments distally bifurcated; elytral surface with digitiform papillae and numerous spinous globular microtubercles from third pair of elytra with macrotubercles amber colour, sclerotized, depressed in honeycomb like groups; outer area with macrotubercles fusiform mucronate, marginal ones hemispherical; inner area with abundant, dark microtubercles conical with thick spines.
Redescription. Holotype of Polynoe brasiliensis (MNHN 78) complete, dehydrated, 1.5 cm long (20–22 mm long in original description), 0.4 cm wide, 26 segments.
Prostomium bilobed, facial tubercle rounded; median antenna with long ceratophore inserted frontally, style dried; lateral antennae inserted terminally with long thin ceratophores, styles dried-out; palps papillate; pharynx not everted.
Tentacular segment not visible dorsally; segment two projected on prostomium as rounded lobe; tentaculophores thin with few thin chaetae.
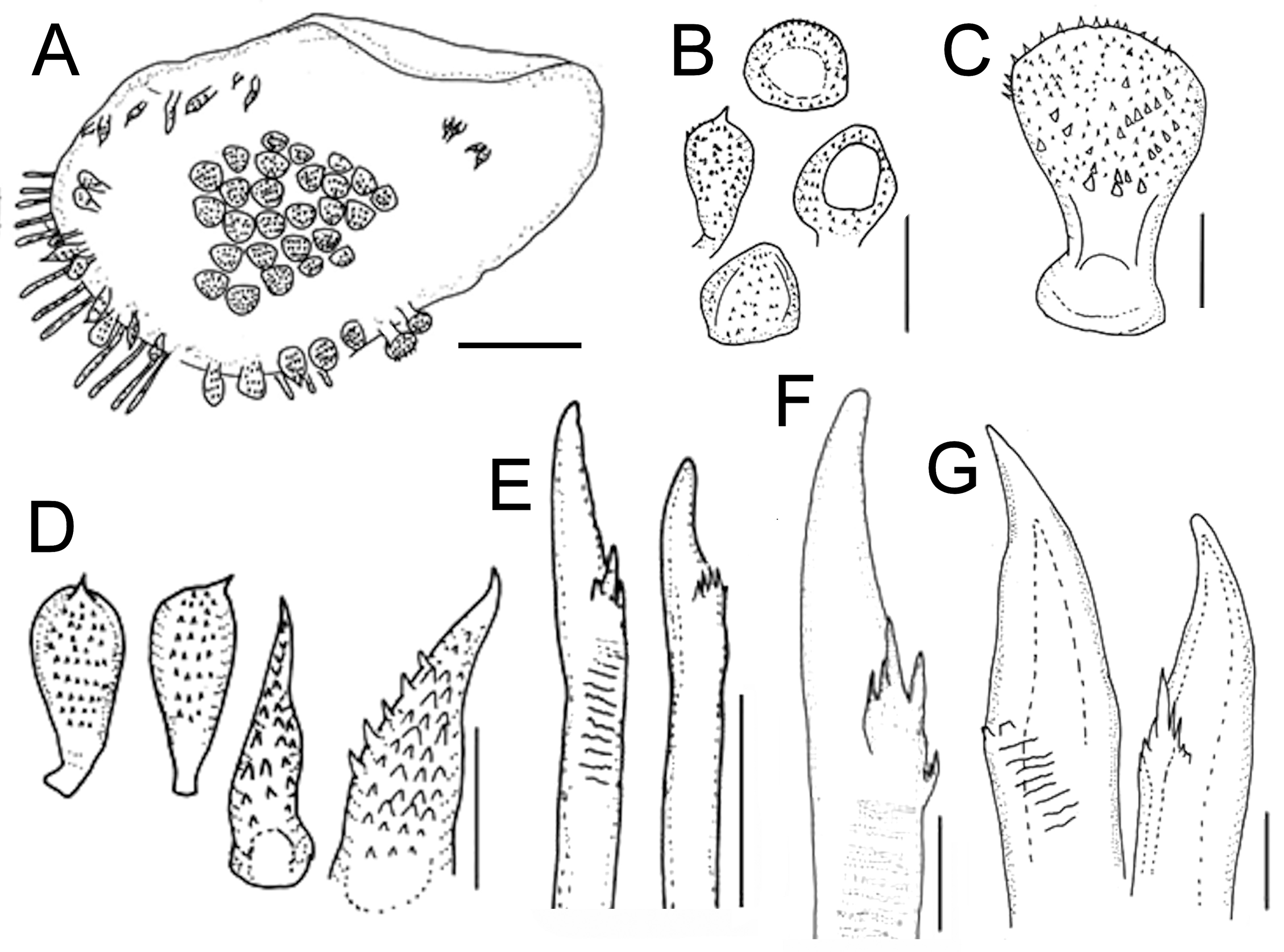
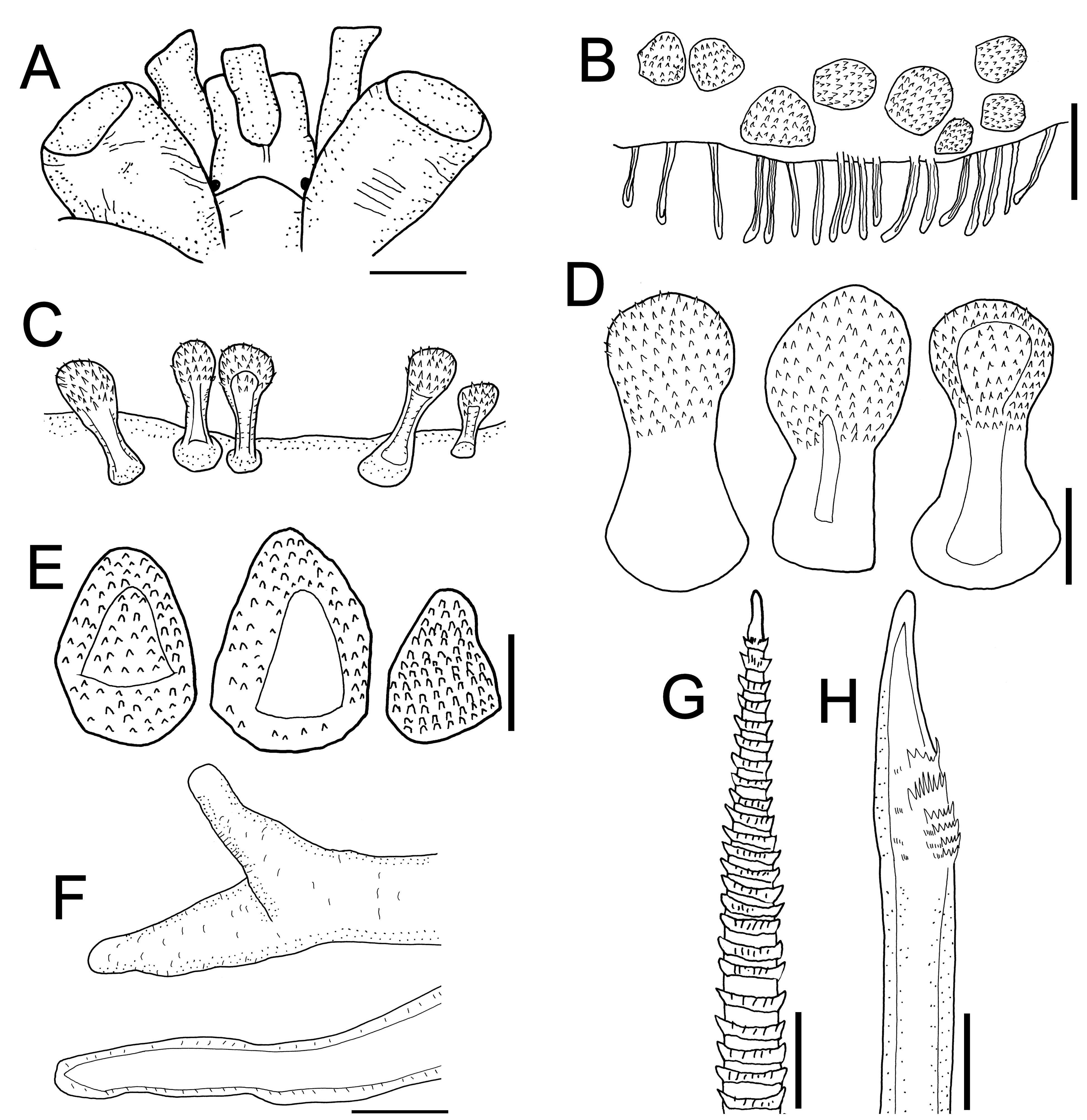
First pair of elytra circular, following ones oval, wide, imbricate, margins with long abundant fimbriae along posterolateral and external margins, and a bundle of long filiform papillae along inner posterolateral margin. Surface of two anterior elytra with hemispherical macrotubercles spinous over elytrophoral area arranged in a C-shaped row. Following elytra with macrotubercles ambercolour, depressed, densely packed in honeycomb like groups in elytral centre ( Fig. 1A View FIGURE 1 ). Outer elytral region with macrotubercles covered by small spines ( Fig. 1B, C View FIGURE 1 ), marginal ones hemispherical, other numerous fusiform with sharp tips, surface spinulose ( Fig. 1D View FIGURE 1 ), markedly larger along posterolateral external area, inner elytral area with abundant microtubercles, conical spinous ( Fig. 1D View FIGURE 1 ).
Parapodia biramous. Notopodia short with abundant notochaetae; neuropodia thick, truncate, with a short acicular projection. Branchial filaments along notopodial anterior and posterior surfaces. Cirrigerous segments with dorsal cirri thin, subdistally swollen, tips long filiform. Ventral cirri thick, short. Notochaetae thin, brownish, shorter than neurochaetae, surface spinulous, tips sharp. Neurochaetae markedly thicker, amber, short, tips unidentate, slightly falcate, with a few subdistal thick spines and striae ( Fig. 1 View FIGURE 1 E–G). Segment 2 with neurochaetae markedly thinner, with several rows of long spines.
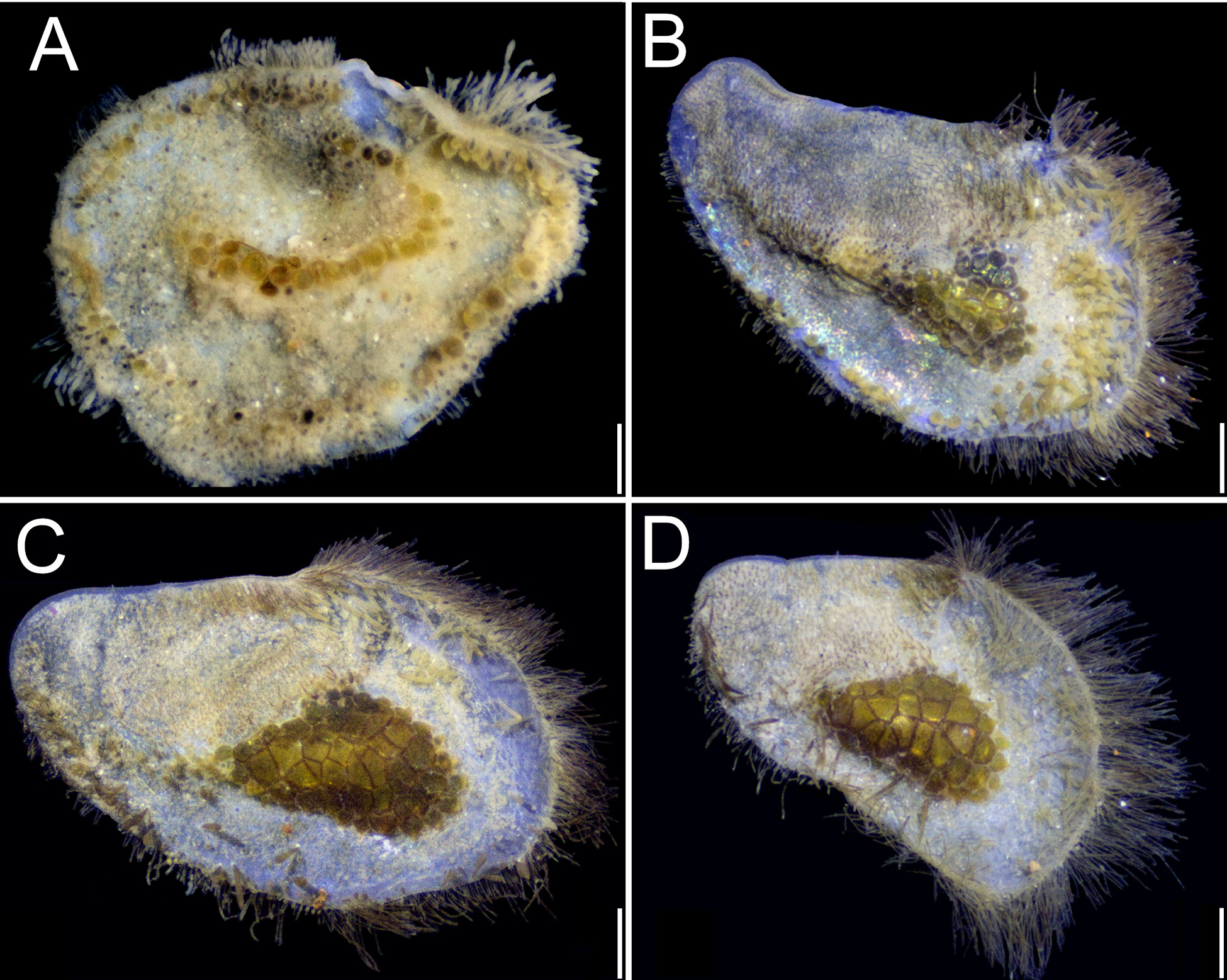
Variation. The holotype of Iphione magnifica Grube, 1876 (ZMB 1059), and the holotype of Polynoe branchiata Treadwell, 1901 (USNM 16006) have the same diagnostic features for prostomia, elytra, and parapodia, with the sole exception that the former does not have its median antenna. Anterior eyes on the widest prostomial area; posterior eyes partially hidden by nuchal lobe ( Figs 2B View FIGURE 2 ; 3B View FIGURE 3 ; 4A View FIGURE 4 ; 6B View FIGURE 6 ). Elytra with fringe of long papillae ( Figs 2C View FIGURE 2 ; 3C View FIGURE 3 ; 5 View FIGURE 5 ; 12 View FIGURE 12 A–C); spinous globular macrotubercles on first elytra ( Figs 2C, D View FIGURE 2 ; 3B, F View FIGURE 3 ; 4B View FIGURE 4 ); from the third pair of elytra macrotubercles are sclerotized, depressed, packed in honeycomb groups ( Figs 2A, E View FIGURE 2 ; 3D View FIGURE 3 ; 4D View FIGURE 4 ; 6A View FIGURE 6 ), with conical microtubercles ( Fig. 3G, H View FIGURE 3 ), and fusiform macrotubercles with acute tips ( Figs 3E View FIGURE 3 ; 5C View FIGURE 5 ). Parapodia with branchial filaments ( Figs 3H View FIGURE 3 ; 5D View FIGURE 5 ; 6A View FIGURE 6 ), and neurochaetae with tips unidentate ( Figs 3I View FIGURE 3 ; 5E View FIGURE 5 ; 6C View FIGURE 6 ). These features are also present in the additional material (UMML 078). Antennae, tentacular and dorsal cirri thin, smooth, subdistally swollen, tips filiform; median antenna longer than prostomium; lateral antennae half as long as prostomium, palps papillated; pygidium thick, long, anal cirri resembling dorsal cirri, subdistally swollen, tips filiform. Elytral ornamentation has been indicated above; the colour of the neurochaetae can vary from light brownish to dark, and the notochaetae are slender and long.
Augener (1927: 44) noted that the elytra of the C. brasiliensis specimens (as Euphione (Chaetacanthus) magnificus ) smaller than 20 mm in length do not show the honeycomb like patch of large macrotubercles. He further added that Treadwell (1901) specimens of P. branchiata were 20–25 mm long, and that because of their small size these patches of elytral macrotubercles were barely developed.
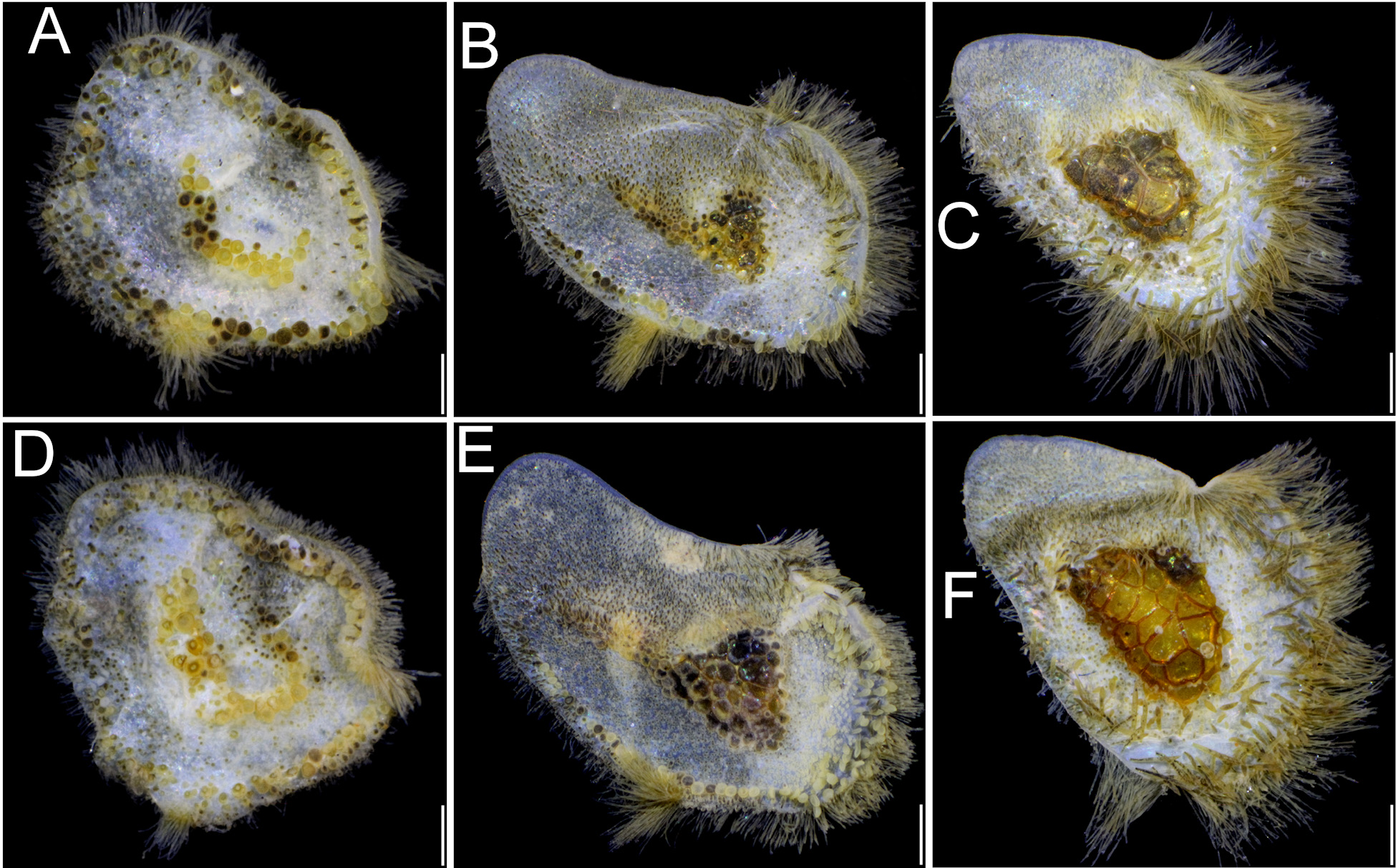
In our new material, elytral shape and ornamentation change along the body. The first elytra are almost circular ( Fig. 7A View FIGURE 7 ); the subsequent ones are elongated, almost oval, medially narrower, and wider laterally, inserted obliquely and imbricate, posterior and lateral margins with fringe. In first and second pair of elytra, the macrotubercles start as an irregular, oblique row ( Fig. 7A View FIGURE 7 ), often with a bent section, resembling a “C”, “J” or half-moon shaped, and there are other large globular macrotubercles along the margin. By segment 9, the macrotubercles are arranged as an ovoid group with individual round and larger macrotubercles ( Fig. 7B View FIGURE 7 ), and the group expands laterally covering about one-fourth of elytral surface ( Fig. 7C View FIGURE 7 ). Elytra of posterior segments show two modifications because macrotubercles become larger although the covered elytral surface is about the same ( Fig. 7D View FIGURE 7 ). Other large macrotubercles are fusiform, truncate basally, distally mucronate, abundant on the outer lateral surface. On the internal elytral areas, the conical spinous microtubercles can be dark. Fimbriae are well-developed along most elytral margins, often loaded with foreign materials, and form a distinct bundle in the inner, posterior elytral region, although they can be eroded by excessive brushing off.
The honeycomb-like group of macrotubercles varies with body size as noted by comparing C. brasiliensis specimens of different size (2.5 cm long vs. 3.5 cm long). The first elytra shows that whereas in smaller specimens ( Fig. 8A View FIGURE 8 ) the central macrotubercles are arranged into a row barely turned around, in larger specimens ( Fig. 8D View FIGURE 8 ), the row turns around quite distinctly resembling an inverted “J”. As indicated above, by segment 9 in smaller specimens the macrotubercles are round, smaller ( Fig. 8B View FIGURE 8 ) than those present in larger specimens ( Fig. 8E View FIGURE 8 ), where they are also polygonal instead of round. In posterior segments, the smaller specimens have more macrotubercles but they are less pigmented, and the group extends over a slightly smaller elytral surface ( Fig. 8C View FIGURE 8 ), whereas in larger specimens, the macrotubercles are larger, darker, and extended over a larger area ( Fig. 8F View FIGURE 8 ).
Remarks. Polynoe brasiliensis de Quatrefages, 1866 is newly combined into Chaetacanthus Seidler, 1922 as C. brasiliensis ( de Quatrefages, 1866) . The original description was based upon a single specimen, hence the holotype, although its type status was regarded as syntype ( Solís-Weiss et al. 2004: S14).
Despite the fact that the original description of P. brasiliensis does not include illustrations, and that the holotype is dehydrated, it is a complete specimen and some diagnostic features were observed: elytral surfaces, and the presence of branchiae are visible between parapodia, close to cirrophores. These features match the diagnosis of Chaetacantus and it explains the new combination. The presence of branchial filaments was overlooked by de Quatrefages (1866), and Seidler (1924) only translated the original description, but the latter transferred the species and newly combined it into Lepidonotus . In that combination, it has been reported in a faunistic note and in a compilation ( Amaral & Nonato 1982, de Assis et al. 2015).
The study of the holotype of Polynoe branchiata Treadwell, 1901 from Puerto Rico, indicated that this species is a junior synonym of C. magnificus as well, as already indicated by Seidler (1924: 97), Augener (1927: 43), and Hartman (1959: 99). We confirm this synonymy, but the senior species name is C. brasiliensis (see below). For instance, the publication date for the paper by Treadwell on Puerto Rican polychaetes has been cited as 1900, 1901, or 1902; however, the correct publication date is 4 October 1901 ( Evermann 1902).
Chaetacanthus brasiliensis and C. magnificus refer to a rare biological entity. It is rarely collected and most collections have just a few specimens, if any at all, with the sole exception of Augener (1927) who reported 30 specimens. Consequently, there are several publications using C. magnificus , and a few ones listing P. brasiliensis . This could make us propose a reversal of precedence and retain the junior synonym instead of the senior one, following article 23.9 of the code (ICZN 1999). This means that because the senior species-group name has been used less frequently than the junior one, the older name should be regarded as a nomen oblitum, and the other more frequently used, nomen protectum. Nevertheless, this violates two essential conditions in 23.9.1.1 and 23.9.1.2 of the Code, because the senior synonym has been used as a valid name after 1899, and the junior synonym has not been used in at least 25 works published by 10 different authors during the last 50 years. Consequently, there is no need to reverse precedence, and we newly combine it as C. brasiliensis , and confirm it as the senior synonym over C. magnificus ( Grube, 1876) .
As explained by Seidler (1924), and herein confirmed, the type specimen of Iphione magnifica Grube, 1876 lacks the median antenna, which probably caused Grube to place it in Iphione Kinberg, 1856 . Despite the fact of this missing appendage, and a superficial similarity in elytral appeareance, the species does not match with Iphione , and it is why Seidler (1924) proposed it as the type species for his new genus Chaetacanthus .
Members of C. brasiliensis resemble C. ornatus n. sp. because their elytra have large macrotubercles in honeycomb-like groups, and a comparison of specimens of similar size indicate some differences ( Fig. 13 View FIGURE 13 ). In C. brasiliensis macrotubercles are smaller, usually polygonal, and extend over smaller elytral areas ( Fig. 13 View FIGURE 13 D–F) than in C. ornatus n. sp. whose macrotubercles are usually larger, globular, and extended over larger elytral areas ( Fig. 13 View FIGURE 13 A–C). On the other hand, Puerto Rican specimens were recorded with ripe oocytes in November ( Allen 1957).
Lepidonotus pilosus Treadwell, 1937 , described from Western Mexico, was also regarded as a junior synonym of C. magnificus by Hartman (1939a: 29; 1956: 253), and by de Brito et al. (2013). However, this is a different species (see below), and it is herein redescribed. On the other hand, Lepidonotus brasiliensis laevis Rullier & Amoreux, 1979 was described with two specimens dredged about 300 km E off Mar del Plata in sediments between 220–270 m depth. The description was restricted to details of elytra and chaetae, and provided illustrations for them. Elytra has large champagne cork-, or mushroom-shaped macrotubercles with finely spinous heads, and long marginal fimbriae; neurochaetae were described and illustrated as blackish, smooth, unidentate. The subspecies was retained in Lepidonotus by de Assis et al. (2015), but its neurochaetae differ from those present in members of this genus and, after this feature, the subspecies does not belong in this genus. Regretfully, branchial filaments were not indicated in the original description, but after Fauchald (1977) key to genera, there would be only two options for transferring this subspecies after the presence of smooth neurochaetae: Euphionella Monro, 1936 if it has branchial filaments, Dilepidonotus Hartman, 1967 if they are missing. For the former, the most similar species might be E. besnardi Amaral & Nonato, 1982 dredged in muddy sands off Rio Grande do Sul, in 200 m depth. For the latter, the only known species, D. falklandicus Hartman, 1967 has blackish unidentate neurochaetae, but macrotubercles are sparser and markedly smaller. Nevertheless, despite Rullier & Amoureux (1979: 152) indicated the type was deposited in the Paris museum (AK 229), it was not included in the type catalogue ( Solís-Weiss et al. 2004). In conclusion, L. brasiliensis laevis does not belong in Lepidonotus nor Chaetacanthus by having smooth neurochaetae; its generic affiliation cannot be solved until the type material is found, or some topotypes are found and carefully compared. This is beyond our current objectives.
Type locality. Bahia, Brazil .
Distribution. Puerto Rico to Panamá, Curaçao, Trinidad, Bahia and Paraíba, Brazil.







| ECOSUR |
El Colegio de la Frontera Sur (Mexico) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Aphroditiformia |
|
Family |
|
|
SubFamily |
Lepidonotinae |
|
Genus |
Chaetacanthus brasiliensis ( de Quatrefages, 1866 )
| Salazar-Silva, Patricia, López-Sánchez, Daniel A. & Salazar-Vallejo, Sergio I. 2020 |
Lepidonotus pilosus:
| Treadwell, A. L. 1941: 26 |
Lepidonotus branchiatus
| Treadwell, A. L. 1939: 184 |
Euphione (Chaetacanthus) magnificus: Augener 1927: 43–44
| Augener, H. 1927: 44 |
Lepidonotus brasiliensis:
| de Assis, J. E. & de Brito, R. J. & Christoffersen, M. L. & de Souza, J. R. B. 2015: 65 |
| Amaral, A. C. Z. & Nonato E. F. 1982: 25 |
| Seidler, H. J. 1924: 37 |
Polynoe branchiata
| Horst, R. 1922: 197 |
Lepidonotus (Physalidonotus) magnificus:
| Augener, H. 1922: 40 |
Chaetacantus magnificus:
| de Brito, R. J. & de Assis, J. E. & Christoffersen, M. L. 2013: 27 |
| Amaral, A. C. Z. & Nonato E. F. 1982: 18 |
| Seidler, H. J. 1924: 97 |
| Seidler, H. J. 1922: 301 |
Physalidonotus magnificus:
| Seidler, H. J. 1921: 87 |
Lepidonotus (Physalidonotus) barbatus
| Augener, H. 1910: 246 |
Iphione magnifica
| Grube, A. E. 1876: 51 |
Polynoe brasiliensis
| de Quatrefages, A. 1866: 247 |