
Ceratocanthus relucens (Bates)
|
publication ID |
https://doi.org/ 10.1649/563 |
|
persistent identifier |
https://treatment.plazi.org/id/03C287A9-9E7E-AB46-5EA4-F6F3FD6950C3 |
|
treatment provided by |
Tatiana |
|
scientific name |
Ceratocanthus relucens (Bates) |
| status |
|
Ceratocanthus relucens (Bates) View in CoL
Third Instar Larva ( Figs. 1–17 View Figs View Figs View Fig ). This description is based on 8 third instar larvae from ‘‘ MÉXICO: Veracruz, Xalapa, Jardín Botánico Francisco J. Clavijero, 1,350 m elevation, 23-V-1999, R . Arce, under pile of old compost’’; 25 third instar larvae, same data except: 26-VII-1999; 12 third instar larvae, 27-IX-1999; 20 third instar larvae, 7-IX-2001. Other 60 third instar larvae collected during May to September 1999 were reared to pupa or adult stage on 20-X-1999 and 22-XI-1999. Specimens are deposited in the collection of Department of Entomology, Instituto de Ecología , A.C. Xalapa, Veracruz, México ( IEXA) .
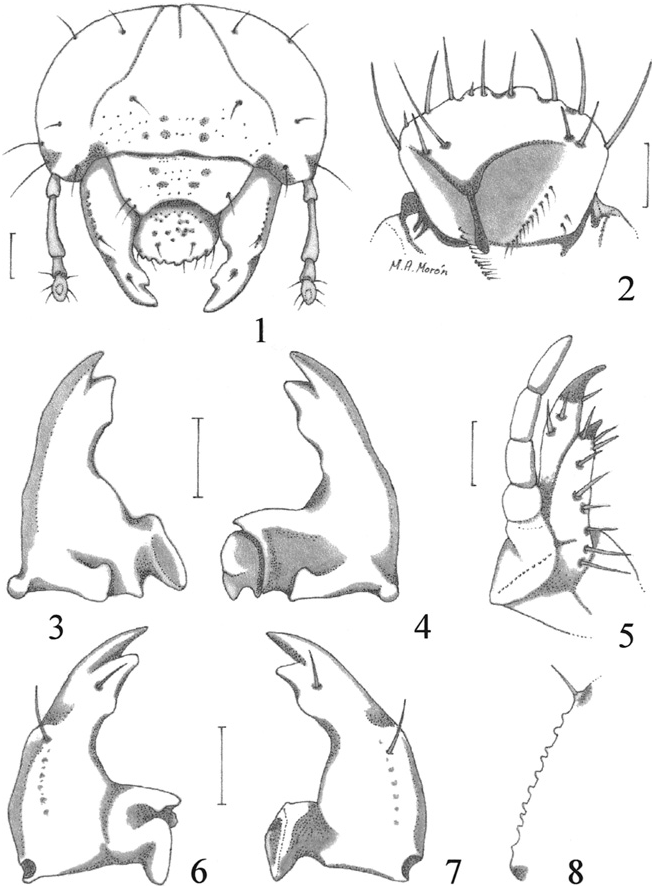
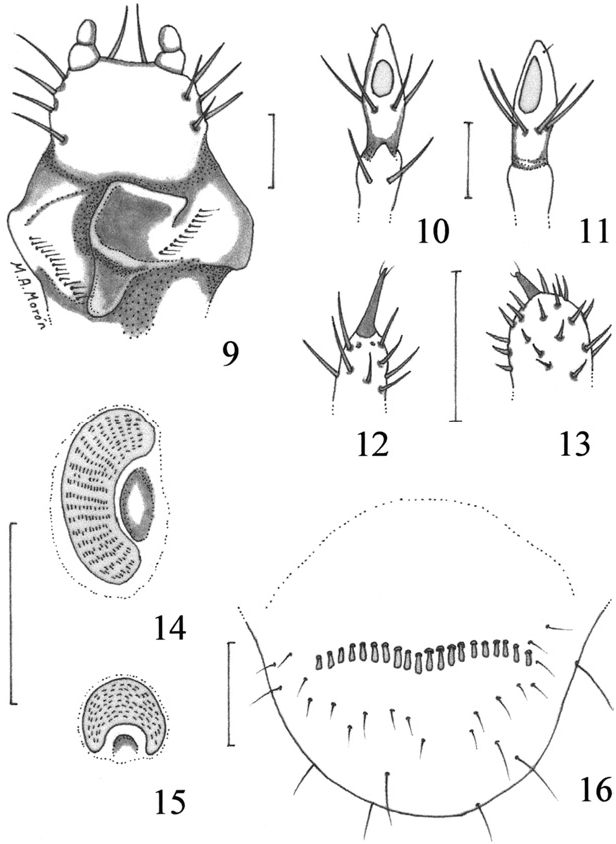
Description. Maximum width of head capsule: 2.1–2.6 mm. Cranium ( Fig. 1 View Figs ). Color light yellow. Surface finely punctate. Frons, on each side, with 1 anterior frontal seta, 1 exterior frontal seta, 1 posterior frontal seta, and 1 seta in anterior angle of frons. Epicranium with 2 dorsoepicranial setae, 1 dorsolateral seta, 2 lateral setae and 1 basi-antenal seta on each side. Epicranial suture complete. Fronto-clypeal suture present, well marked. Stemmata absent. Clypeus. Asymmetrical, produced at the anterior right angle, with 1 seta at each side, and 2 anterior clypeal setae. Labrum. Asymmetrical, with shallow, sparse punctures, 2 anterior labral setae, 2 setae near the basal border, 1 setae at each side, and anterior margin with 4 truncate teeth. Epipharynx. ( Fig. 2 View Figs ), with a dextral beak-like process, paria without chaetae. Left paria with a laeophoba of 9–11 filaments; right paria with a dexiophoba of 9–12 filaments toward the basis. Tormae united; anterior epitorma wide and complete; posterior epitorma short, rounded; right pternotorma bifurcate; left pternotorma long, acute. Mandibles. Asymmetrical. Rigth mandible ( Figs. 3, 7 View Figs ) with 1 apical, acute scissorial tooth followed by 1 wide, scissorial blade. Without stridulatory area. Molar area wide, pedunculate. Brustia absent. Left mandible ( Figs. 4, 6 View Figs ) with 1 apical, acute scissorial tooth and 1 wide scissorial blade, with the border widely sinuated. Without stridulatory area. Molar area complex, bilobed, apical molar lobe, short, transverse and depressed, basal molar lobe wide, dorso-longitudinally compressed. Brustia absent. Maxilla. ( Fig. 5 View Figs ) with separate galea and lacinia; uncus of galea much longer and wider than the uncus of lacinia; stridulatory area ( Fig. 8 View Figs ) consisting of a row of 10 rounded, some vaguely conical teeth and a distal, tubercle-like process with short, slender seta. Labium. ( Fig. 9 View Figs ). Glossa smooth, with 4–5 setae at each side. Hypopharyngeal sclerome asymmetrical, with upturned, anterior, sclerotized process, without setae at sides. Labial palpi formed by 2 segments; last labial segment slightly longer than wide, with rounded apex. Antennae. 4-segmented, with second segment more than three times longer than first segment, and longer than the lenght of the combined third and fourth segments. Last antennal segment not reduced in diameter, with 1 dorsal and 1 ventral sensory spots, 6 slender, long setae around the middle, and 1 diminute preapical setae; third antennal segment without sensoria ( Figs. 10–11 View Figs ).
Thorax. ( Fig. 17 View Fig ). Pro- meso- and metanota without sclerotized lateral areas. Scutella of meso- and metathorax bare. Thoracic spiracles ( Fig. 14 View Figs ) with concavity of respiratory plate facing posteriorly; spiracle closing apparatus absent; size 0.20 mm high and 0.06 mm wide; distance between the two lobes of the respiratory plate very much wider than the dorsoventral diameter of the bulla; plate with 10–14 holes across diameter at middle, holes elongate, arranged in transverse rows. Bulla of spiracle slightly convex.
Legs. ( Fig. 17 View Fig ). With minute, oblique, truncate stridulatory carinae on the external side of mesofemur (discernible only with a compound microscope); pro- and metafemora without such structures. Each tarsal claw bearing 2 short apical setae; metatarsal claws clearly shortened, with rounded apex ( Figs. 12–13 View Figs ).
Abdomen. Abdominal spiracles ( Fig. 15 View Figs ) with concavity of respiratory plate facing ventrally. Spiracle closing apparatus absent. Spiracle of 1st abdominal segment 0.10–0.11 mm high and 0.05–0.06 mm wide; spiracles of 2nd to 7th abdominal segments 0.08–0.09 mm high and 0.02– 0.03 mm wide; spiracle of 8th abdominal segment 0.08–0.10 mm high and 0.04–0.05 mm wide. Dorsa of 1st to 7th and 9th and 10th abdominal segments each with 1–3 transverse rows of scattered, medium size, slender setae. Dorsum of 8th abdominal segment with transverse row of 4–6 slender, long setae, near the anterior border. Venter of last abdominal segment ( Fig. 16 View Figs ) with transverse palidia of 10–18 spatulate pali. Anal slit transverse, located toward the dorsal surface of last abdominal segment ( Fig. 17 View Fig ).
Remarks. Third instar larva of C. relucens may be distinguished from Germarostes larvae mainly by the dorsal surface of labrum scarcely and irregularly punctate, without coarse tubercles or strong keels. The anterior margin of the labrum usually presents four truncate teeth (76% of observed specimens), but some larvae have three or five teeth, including an abraded lateral tooth, or by a briefly notched central tooth. The number of pali varies from eight to 18, but more commonly the palidia have 13–14 pali (43% of observed specimens).
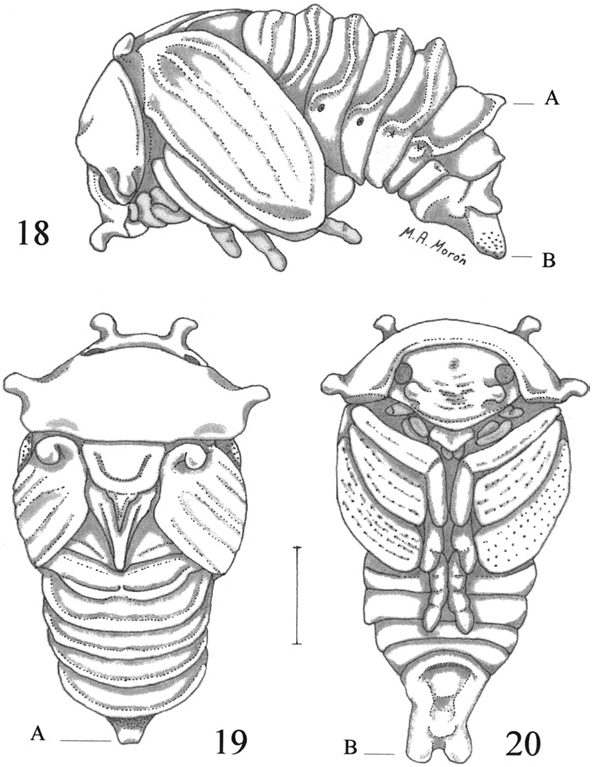
Pupa. ( Figs. 18–20 View Figs ). This description is based on 12 pupae from ‘‘ MÉXICO: Veracruz, Xalapa, Jardín Botánico Francisco J. Clavijero, 1,350 m elevation, 21-X- 1999, R . Arce, under pile of old compost.’’ Specimens are deposited in the collection of Department of Entomology of Instituto de Ecología, A.C. Xalapa, Veracruz, México ( IEXA) .
Description. Body elongate, robust, exarate ( Fig. 18 View Figs ). White yellowish. Without velvety microtrichia on the abdominal segments. Head. Reflexed downward; antennae and mouthparts clearly separated; ocular canthus and compound eyes well differentiated; surface of frons slightly convex; clypeus convex, slightly rugose, with one clavate projection, apically rounded, near each ocular canthus; labrum wide, flattened ( Figs. 18–20 View Figs ).
Thorax. Pronotal disk convex, with shallow depressions toward the sides; lateral margins defined, and with one clavate projection, apically rounded, at each side ( Figs. 19–20 View Figs ). Meso- and metanota differentiated. Meso- and metascutellum narrowed backward, apex acute or nearly rounded ( Fig. 19 View Figs ). Pteroteca free, but closely compressed around body; hind wing teca nealy as long as the elytron teca; each elytron with one clavate projection, apically rounded, near the humeral area ( Figs. 18–19 View Figs ). Meso-metasternal areas covered by legs. Protibia without projections on external border; meso- and metatibia widened, striated, without projections on external or apical borders; all tarsomeres vaguely defined.
Abdomen. Tergites without dioneiform organs; 1st tergite with a rounded, transverse carina near the middle; 2nd to 5th tergites each with a strong, sinuose, transverse carina; 6th tergite with strong transverse carina toward posterior margin, bearig a noticeable, truncate projection at middle ( Figs. 18–19 View Figs , point A); 7th tergite convex, with a rounded, brief, transverse carina near the posterior margin; 8th tergite with a wide, transverse depression. Tergo-lateral tubercles fused with tergal carinae. Spiracle of 1st segment elongate, hidden under posterodorsal fleshy folds and hind wing teca. Spiracles of 2nd to 4th segments with ovate, ring-like, sclerotized peritreme and open atrium. Spiracles of 5th to 7th segments with atrium closed, without peritreme. Sternites of 2nd to 7th segments convex. Last segment with 2 rounded, fleshy projections on the posterior border, probably urogomphi, without sclerotized spines ( Figs. 18, 20 View Figs , point B). Genital ampulla short, narrowed, slightly convex.
Remarks. The pupa of C. relucens has three pairs of clavate projections on the clypeus, pronotum and elytra, and the urogomphi do not have sclerotized spines at the apex. The pupa of G. macleayi described by Costa et al. (1988) has only one pair of clavate projections near the base of the elytra, and the urogomphi have sclerotized, acute spines at apex. The pupa of C. aeneus illustrated by Choate (1987) also present three pairs of clavate projections, but Choate said that these structures are located ‘‘around the head and thorax (two on front of head, four on thorax).’’ The lateral abdominal profile of the pupa of C. aeneus photographed by Choate is similar to the pupa of C. relucens , but the details of the urogomphi are not clear.
Biological Notes. Second instar larvae are very similar to third instar except in size, with maximum width of head capsule: 1.2–1.3 mm. The observed old compost pile offered a good site for the development of C. relucens , because we found specimens of different instars through the Summer, Autumm and early Winter spanning a one year period. During May we found third instar larvae; during June and July third instar larvae, pupa and teneral adults; during August third instar larvae, pupae and adults were collected; during September, first, second and third instar larvae, some immature adults and many mature adults were obtained; during October, some pupae, teneral and mature adults were found; during November, only adults were collected inside pupal cells; and during December and January, third instar larvae and mature adults were obtained. Between February and April, we did not collect in the compost. During January, May and July we obtained adult specimens attracted to white fluorescent light in the same area. Apparently, life cycles of different generations are partially overlapped in this locality, but flying adults during May–July and January suggest the development of two generations each year.
With the specimens reared in the laboratory, we obtained the following data: third instar larvae feed for 45–52 days; the completely developed larva builds an oval pupal chamber (8–10 mm in diameter) with excrement and surrounding particles. The walls are thick and very hard once they are completely dried. The prepupa stage does not feed and reqires 30 days, and pupation requires 24–32 days. These data suggest that the complete life cycle requires nearly 180 days, and supports the hypothesis of a bivoltine condition for this population.
Choate (1987) observed that during May the larvae of C. aeneus spent two weeks in pupal cells before pupation. Pupae required approximately two weeks to emerge as adults; teneral adults remained in pupal cells at least one week prior to emergence during June. But, Choate also observed teneral adults and larvae in the breeding site during other months, such as February and March.

| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.