
Centropus maximus, Shute & Prideaux & Worthy, 2016
|
publication ID |
https://doi.org/ 10.1111/zoj.12387 |
|
publication LSID |
lsid:zoobank.org:pub:A76E15BF-087D-43AB-8D53-BF40349561E2 |
|
persistent identifier |
https://treatment.plazi.org/id/F2404834-F927-489E-B5E3-9A79A602CB24 |
|
taxon LSID |
lsid:zoobank.org:act:F2404834-F927-489E-B5E3-9A79A602CB24 |
|
treatment provided by |
Marcus |
|
scientific name |
Centropus maximus |
| status |
sp. nov. |
CENTROPUS MAXIMUS SP. NOV. ( FIGS 2, 3 AND 5)
urn:lsid:zoobank.org:act:F2404834-F927-489E-B5E3-9A79A602CB24
Holotype ( Figs 2, 3 and 5)
WAM 09.3.283: associated remains of one adult individual comprising: fragments L and R mandibular rami; R pterygoid; nine vertebrae (vert.) assessed against Ce. violaceus (cervical vert. 3, 4, and?6; cervicothoracic vert. 12; thoracic vert. 14, 15, 16, 17, and 18); seven ribs and rib fragments; R, pL, dL humerus; pL, dL ulna; L coracoid; omal fragment L clavicula; pL and part of corpus, mid-portion of R corpus, scapula; cranial portion of sternum, including cranial end of carina; pelvis, missing ala postacetabularis ilii, and most of both pubi and ischii; dL, R femur; fragment of pL tibiotarsus; pL tarsometatarsus; various fragments; NISP = 35. Measurements of long bones are given in Table 3; additional measurements are given in text where applicable.
Paratype
WAM 09.3.284: associated remains of one adult individual comprising: cranium, five fragments; L pars caudalis and tip mandible; L missing processus mandibularis, R quadrate; fifteen vertebrae (vert.) (cervical vert. 5-13; thoracic vert. 14-18; one caudal vert.); L, R humeri; R ulna missing olecranon process; R carpometacarpus, missing os metacarpale minus; R phalanx digiti majoris; L coracoid, missing omal portion; pL and part of corpus, scapula; cranial fragment sternum; part furcula; pR, pL, dL femora; L, R tibiotarsi; R fibula; L, dR tarsometatarsi; five L pedal phalanges, (I.1; III.1; III.2; ungual?III.3; IV.1); NISP = 47. Measurements of long bones are given in Table 3; additional measurements are given in text where applicable.
Type locality
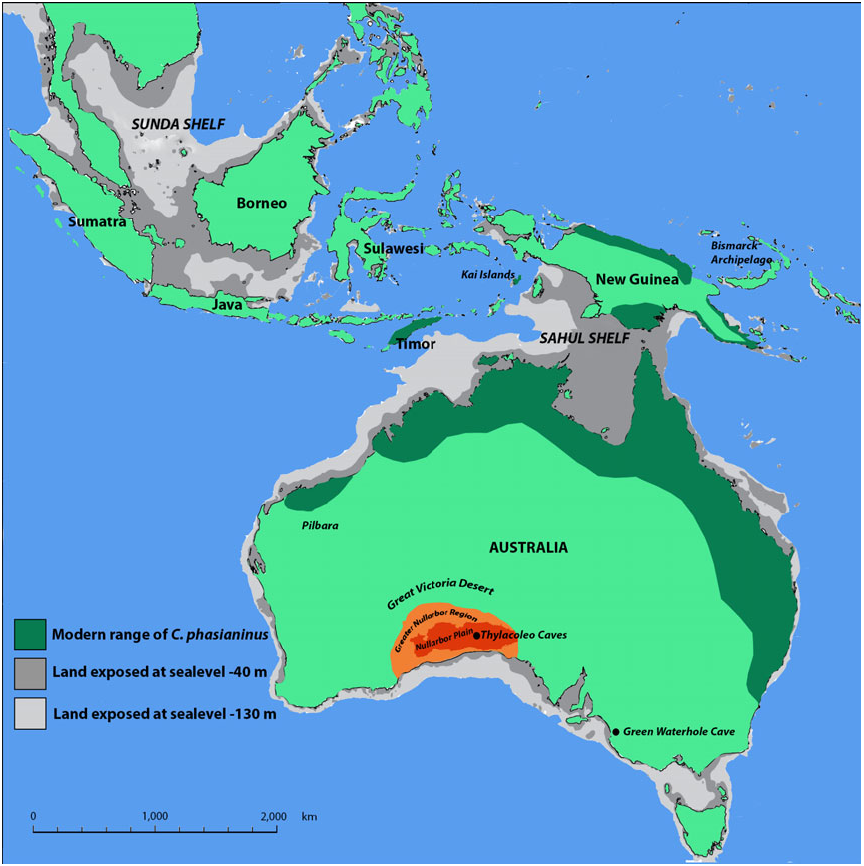
Flightstar Cave, Nullarbor Plain, Western Australia ( Fig. 1 View Figure 1 ). The precise location for the site is registered with the Department of Earth and Planetary Sciences, Western Australian Museum, Perth. Flightstar Cave is one of three caves comprising the Thylacoleo Caves. It is formed within the Early Miocene-aged Nullarbor limestone and preserves a vertebrate fossil assemblage that is of Early to Middle Pleistocene age ( Prideaux et al., 2007).
Stratigraphy, age, and fauna
The holotype and paratype were shallowly buried in powdery limestone breakdown material between boulders in the rockpile beneath the solution-pipe entrance, through which the specimens of Ce. maximus are presumed to have accumulated via pitfall trapping. The femur (09.3.283a) of the holotype was collected on 30 April 2014 by Lindsay Hatcher and Michael Simpson, and the remainder of the holotype and all bones of the paratype were collected by them the following day (1 May). Upper infill sediments elsewhere in the cave, upon which other extinct megafauna were found, have a minimum age of 230 ƚ 27 kyr ( Prideaux et al., 2007). The holotype and paratype of Ce. maximus are presumed to belong to the same fossil fauna, and thus are probably of Early or Middle Pleistocene age. The holotype and paratype skeletons were found adjacent to one another and have similar preservation, and are likely to have died at the same time.
Diagnosis
The largest known extinct or extant species of Centropus , with a humerus slightly longer than that of Ce. colossus , but with a proportionally more slender shaft, a wider and more robust proximal end, a less inflated caput humeri that does not extend further proximally than the tuberculum ventrale in ventral aspect, a strongly marked scar for the insertion of the m. supracoracoideus between the caput humeri and the crista deltopectoralis orientated proximodorsally, a more inflated intumescentia humeri, a deeper impressio coracobrachialis, a more prominent impressio m. pectoralis, a crista bicipitalis that is very robust and craniocaudally thick distally, a shaft that is not ventrally inflated around the fossa m. brachialis, a more prominent processus supracondylaris dorsalis, a more robust processus flexorius that is as deep as it is long distad of the tuberositas supercondylare ventrale, and with larger, more robust leg bones.
The humerus of Ce. maximus is considerably larger than those of any extant species of Centropus , including the large Melanesian taxa Ce. milo and Ce. violaceus , indicating that this species was of much greater size than any living coucal. It is also much larger than the humerus of the extinct Australian Pleistocene species Ce. bairdi sp. nov. described above. Based on published tarsus lengths of cuculids ( Erritzøe et al., 2012), the tarsometatarsus of Ce. maximus (measured without the eminentia intercondylaris as it would be measured in live birds and preserved skins) is, at 112.9 mm, also nearly 25 mm longer than that of the next largest cuculid ( Carpococcyx renauldi = 89 mm; Erritzøe et al., 2012).
Etymology
Maximus = ‘largest’ (Latin, masculine), to acknowledge the very large size of this species, which was greater than that of any known member of the genus, and indeed greater than any known cuckoo species.
Description and comparisons
The bones of the holotype and paratype are off-white in colour, stained slightly in places by orange/brown sediment. They are very fragile and some are damaged, with breaks (e.g. shaft breakages of long bones) having occurred to some bones during collection. Where possible, breaks have been repaired. Although damaged in places, the bones are not significantly eroded, and preserve anatomical detail very well. The bone surfaces lack a porous texture and the epiphyses of the tarsometatarsi of both individuals are completely fused; therefore, both are fully grown adult birds, the paratype being a slightly larger individual.
Cranial bones: The mandibular fragments show that in comparison with Ce. violaceus , the symphyseal area is proportionally shorter and shallower, and is not drawn out into a deep, narrow, elongate, downcurved tip: the shape is more like that of Ce. phasianinus . The caudal dentary fragments of WAM 09.3.283 (L and R) preserve the area transitioning from covered to uncovered in rhamphotheca in modern specimens. At 12.2 mm high at the caudal end of the dentary, the bill was some 20% bigger than that of Ce. violaceus . The pars caudalis of the mandible lacks the processus medialis but the preserved parts are very similar to those of the species of Centropus examined, and overall the pars caudalis is 30% larger than that of Ce. violaceus . The pterygoid is 19.3 mm long and essentially the same shape as in Ce. violaceus . The R quadrate is well preserved, and at 22 mm high, some 22% taller than that of Ce. violaceus (18 mm). It is larger overall than that of Ce. violaceus and differs from it only by the presence of a large foramen pneumaticum caudomediale sensu Elzanowski & Stidham (2010), by lacking a foramen pneumaticum rostromediale (a small hole in this facies is interpreted as damage), and by having a pars quadratojugalis and fovea quadratojugalis that are less prominent than in Ce. violaceus , much less so than in Ce. phasianinus .
Humerus: As well as the generic characters described above, the humerus of Ce. maximus has the following features: a thin crus dorsale fossa; a large and deep impressio coracobrachialis occupying nearly half of proximal width; the contour of the crista bicipitalis is reduced distoventrally; the impressio m. pectoralis is about 10 mm long and noticeably elevated off the adjacent facies distally; a thick crista deltopectoralis; and a low caput humeri as in Ce. violaceus .
Ulna: WAM 09.3.284 is nearly complete, with a little damage at the proximal and distal ends. The olecranon is damaged, so the total length of the bone is probably about 1 mm more than measured if shaped like in Ce. violaceus . The ulna is distinguished from those of all extant Australian cuculids, except for S. novaehollandiae , by its considerably greater length and broader shaft ( S. novaehollandiae has an ulna around 15% longer). The ulna is distinguished from that of S. novaehollandiae by having: a strongly dorsoventrally flattened shaft, rather than being round in crosssection; a very reduced and poorly marked impressio brachialis; a less distally extended processus cotylaris dorsalis; a smaller, less projecting tuberculum carpale; and extremely large and protuberant papillae remigalis caudalis. The ulna of Ce. maximus , although larger, is similar in shape to those of Ce. phasianinus, Ce. bengalensis , and Ce. violaceus . Proportionally it is most similar to that of the near-flightless Ce. violaceus than to those of the more volant species, having a robust, especially flattened shaft that widens considerably as it approaches the proximal end and a similarly weakly marked impressio brachialis. However, it is distinguished from the ulna of Ce. violaceus by its L, left; R, right; TL, total length; dL, distal left; dR, distal right; pL, proximal left; pR, proximal right; PW, proximal width; SW, mid-shaft width; DW, distal width; *, minimum measurement owing to damage; †, measurements of coracoid are as follows: TL is measured medially, PW is the width of the omal end, DW is the width of the sternal end; §, proximal width of scapula is measured from the acromion to the ventrodistal margin of the facies articularis humeralis.
larger size, its much larger papillae remigalis caudalis, and a cotyla ventralis that is proportionally wider dorsoventrally. The ulna of Ce. colossus is not known, but based on humeral/femoral proportions may be of similar size to that of Ce. maximus . The ulna of Ce. bairdi sp. nov. is also unknown, but is presumed to be considerably smaller than that of Ce. maximus .
Carpometacarpus: WAM 09.3.284-k (L) preserves the entire length but lacks the os metacarpale minus, the proximoventral portion of the bone is damaged, and the processus pisiformis and surrounding area are missing. Morphologically it is very similar to, but about 25% longer, and more robust, than those of the extant Ce. phasianinus and Ce. violaceus , and is much bigger than that of Ce. bengalensis . It is further distinguished from the carpometacarpus of Ce. violaceus by having subequal distal extension of the facies articularis digitalis major and the facies articularis digiti minoris, whereas in Ce. violaceus the facies articularis digiti minoris projects rather more distally. The sulcus tendineus is deeper and the shaft in cranial view is straighter (proximally it is tilted ventrally in Ce. violaceus ).
Coracoid: WAM 09.3.283 (L) is near complete, missing only the tip of the processus procoracoideus and the lateral margin of the processus lateralis. It is distinguished from the coracoid of all large extant Australian cuculids and that of the Melanesian Ce. violaceus by its much greater size and its proportionally stouter shaft. The omal width is proportionally wide and the facies articularis clavicularis is large as in Ce. phasianinus and Ce. bengalensis , whereas Ce. violaceus is narrower here and has a reduced facies articularis clavicularis. The processus procoracoideus is slightly damaged at the tip, but is proportionally narrower than that of other Centropus species. The lateral extent of the processus lateralis cannot be determined. The acrocoracoid is not penetrated by pneumatic foramina from the sulcus supracoracoideus beside the facies articularis clavicularis, as in Ce. violaceus but unlike in Ce. phasianinus , which is pneumatic. Also in ventral view, the sternal margin of the sulcus supracoracoideus/acrocoracoid is only slightly cranial to the sternal margin of the facies articularis humeralis.
Scapula: WAM 09.3.283 (L) is the most complete specimen, preserving the cranial end but lacking the caudal extremity, whereas 09.3.283 (R) lacks its cranial end and caudal extremity but preserves most of the shaft. The scapula is distinguished from those of all extant Australian cuculids and that of the Melanesian Ce. violaceus by the greater size of the articular surfaces and the width of the corpus. Although neither scapula from the holotype preserves the entire length, when both are aligned side by side, the estimated total length of the scapula of this individual is> 64 mm. It has pneumatic foramina medially caudal to the tuberculum coracoideum, which is only a small medial prominence. The acromion is robust with a prominent laterally directed facies articularis clavicularis.
Sternum: WAM 09.3.283 (holotype) has the most complete specimen, preserving around 1.5 cm of the length of the cranial portion of the sternum, including the sulci articularis coracoidei, L and R processus craniolaterales, and the full anterior depth, although not the length, of the carina. Amongst the sterna of the species of Centropus that were available to us, it bears closest resemblance to Ce. violaceus in both size and morphology, being somewhat larger and more robust, and sharing its lack of a spina externa (see phylogenetics results), and having a very shallow, thick keel that lacks a cranially projecting apex carinae (carina relatively deeper, thinner, and with a cranially projecting apex carinae in Ce. bengalensis and Ce. phasianinus ). Besides its greater size and robusticity, the sternum of Ce. maximus is distinguished from that of Ce. violaceus by the following features: the profile is less dorsoventrally flattened in cranial aspect so that the sternal basin is relatively deeper; the carina is somewhat proportionally deeper; the labrum externa of the sulcus articularis coracoideus projects further cranially either side of the midline (more or less flat in Ce. violaceus ); and there is a distinct gap between the left and right sulci articularis coracoidei (they form a continuous groove in Ce. violaceus ). The overall size of the cranial portion of the sternum is similar to that of S. novaehollandiae , but Ce. maximus is distinguished from this species by being much more robust with thicker bone, having a much shorter and thicker carina that does not project as far cranially as the labrum externa of the sulcus articularis coracoideus, and lacking a spina externa (thin and prominent in S. novaehollandiae ). This specimen preserves four processus costales and the processus craniolaterales are cranially directed as in other species of Centropus . The carina of this species is reduced relative to that of Ce. phasianinus (approx. 17% of femur length in Ce. maximus vs. 23% in Ce. phasianinus ), but less reduced than in Ce. violaceus (approx. 12%), suggesting that the extinct taxon may have been more volant than Ce. violaceus . Measurements (mm) ( WAM 09.3.283): cranial width,> 42.9; depth of carina, 18.0; maximum width of carina, 5.5.
Pelvis: WAM 09.3.283 (holotype) preserves the only known pelvis. It reveals the complete length of the synsacrum, but neither left nor right ala postacetabularis ilii, ala ischii, nor pubii survives. The pelvis is distinguished from that of all large extant Australian cuculids and that of Ce. violaceus by its much greater size and robusticity. Compared to pelves of all other species of Centropus examined, including Ce. violaceus , the cranial end is absolutely and proportionally much deeper dorsoventrally, and the foramen acetabuli is absolutely and proportionally much larger. The ala preacetabularis ilii is very deeply concave, creating a large fossa iliaca dorsalis. Owing to damage to the bone surface, it is not possible to determine if the ala preacetabularis ilii is perforated as in Ce. bengalensis, Ce. phasianinus , and Ce. ateralbus , or solid as in Ce. violaceus , but two holes are present in the relevant place and at least attest to the ilium being very thin at this point, even if it were not perforated before death. If the cranially projecting process at the cranial extreme of the dorsal surface was present as in other species of Centropus , it is not preserved in this specimen, but, as preserved, the left side indicates that Ce. maximus had a less acute proximal extension to the fossa iliaca dorsalis. Measurements (mm) ( WAM 09.3.283): length of synsacrum, 85.9; maximum width (measured between the caudal rims of the left and right antitrochanters), 60.9; minimum width (measured at the narrowest part of the alae preacetabulares ilii), 23.8; cranial depth, 29.9.
Femur: WAM 09.3.283 (R) is the most informative specimen, and is complete with some damage to the facies articularis acetabularis on the caput, as well as to the condylus medialis and the cristae lateralis and medialis sulci patellaris. It is very large and robust, and is far larger than the femur of any extant Australian cuculid, including Ce. phasianinus , and is also considerably larger than the femur of the extinct Ce. bairdi sp. nov. described above, and that of the Melanesian taxon Ce. violaceus . It is similar in length and shaft width to the femur of Ce. colossus described above, but is slightly longer and is more robust at its proximal and distal ends. It is distinguished from the femur of Ce. colossus by the following features: the collum femoris is craniocaudally thicker; the facies articularis antitrochanterica projects caudally, and the caudal facies distal of this projection is convex, as in Ce. bairdi sp. nov., rather than concave as in Ce. colossus ( Fig. 3); on the craniomedial facies between the trochanter and the collum femoris there is a marked insertion for the m. iliofemoralis internus, which has about half the area of that in Ce. colossus ; the impressiones obturatoriae interna and externa are larger and deeper; the shaft is craniocaudally thicker where it meets the proximal end of the bone, and the trochanter femoris is wider in lateral aspect; the distal end has wider and deeper condyles; the trochlea fibularis is deeper and wider, and is separated from the fossa poplitea by a zone containing a shallow fossa, and a proximocranial extension of the crista tibiofibularis; the impressio ligamentum cruciati cranialis is larger and deeper; the fovea tendineus m. tibialis cranialis is larger and deeper; and the origin of the ligamentum cruciati caudalis is smaller, less prominent, and is more medially located on the lateral wall of the fossa poplitea. Measurements (mm): for TL, PW, SW, and DW see Table 3; minimum shaft circumference = 26.2.
Tibiotarsus: WAM 09.3.284 (L and R) show that the tibiotarsus of this species is remarkably large and robust, roughly double the length of the tibiotarsus of its extant Australian congener, Ce. phasianinus . The tibiotarsus of Ce. colossus is not known, but given that its distal femur is less robust than that of Ce. maximus (see above), its tibiotarsus would also be expected to be slightly more gracile. The tibiotarsus of Ce. maximus is markedly larger (nearly 30% longer) than that of the extant Ce. violaceus and the extinct Ce. bairdi sp. nov. described above. Apart from its larger size, the tibiotarsus is further distinguished from those of Ce. bairdi and Ce. violaceus by the following features: the facies gastrocnemialis is even more flattened than in Ce. bairdi , but the flattened surface of the shaft distal of the facies only extends about one-fifth of the length along the medial facies of the shaft, rather than half way; there is a deep depression at the proximocaudal part of the facies gastrocnemialis; the foramen interosseum proximale is very deep and crescent-shaped, and is readily apparent even in the absence of an articulated fibula; the pons supratendineus is about twice as wide as it is proximodistally long, and proportionally shorter proximodistally than in Ce. bairdi ; the tuberositas retinaculi extensori for the insertions of the transverse ligament are very distinct (less pronounced in Ce. bairdi and even less so in Ce. violaceus ), and the proximal tuberosity is relatively more distally located (proximal distance from pons equivalent to only half the shaft width at the pons) than in Ce. bairdi and even more so than in Ce. violaceus (proximally, distance from the pons is equivalent to shaft width at the pons); the sulcus extensorius is more laterally located than in Ce. bairdi , and especially so relative to Ce. violaceus ; the incisura intercondylaris is wider than in Ce. violaceus and the condyles diverge widely cranially (parallel in Ce. violaceus ), more so than in Ce. bairdi ; and the condylus medialis is markedly deeper than it is proximodistally long (round in Ce. violaceus , off round in Ce. bairdi ). Measurements (mm): for TL, PW, SW, and DW see Table 3. WAM 09.3.284d: width at crista fibularis,> 16.1; depth lateral cotyla, 15.0; depth medial cotyla, 15.7; minimum shaft circumference = 23.6.
Tarsometatarsus: WAM 09.3.284-f is the most informative specimen, preserving the entire length of the bone but with damage to the hypotarsus and trochleae metatarsi II and IV, whereas WAM 09.3.284-g preserves trochlea metatarsi II. The tarsometatarsus of Ce. maximus is much larger than that of any extant cuculid species, and is also considerably larger than that of the extinct Ce. bairdi described above. The tarsometatarsus of Ce. colossus is not known, but was probably slightly smaller than that of Ce. maximus , given its slightly smaller and more gracile femur. In addition to the features mentioned in the generic diagnosis for Centropus above, the tarsometatarsus of Ce. maximus has the following features: the tuberculum m. fibularis brevis is small and projects laterally at the lateroplantar corner of the lateral cotyla, as in Ce. bairdi ; the impressiones retinaculi extensorii are low and small; the foramen vascularis medialis is placed more proximally than the foramen vascularis lateralis, but both are of similar size; the tuberositas musculus tibialis cranialis is very large, is placed on the medial wall of the sulcus extensorius, and there is a distinct gap between it and the foramen vascularis medialis (see phylogenetics results); the sulcus flexorius is deeply excavated from the fossae parahypotarsales to almost as far distal as fossa metatarsi I; fossa metatarsi I is very deep, but does not excavate the medial shaft margin; trochlea metatarsi III is wider than it is dorsoplantarly deep, and its dorsal and plantar widths are the same (i.e. the planes of the trochlea are parallel, and do not converge plantarly as in Ce. violaceus ) and it has a broad, deep medial groove; and the medial surface of the shaft is thick as in Ce. bairdi , unlike the dorsoplantarly compressed margin seen in Ce. violaceus . Measurements (mm): for TL, PW, SW, and DW, see Table 3. Proximal depth incl. hypotarsus: WAM 09.3.284 (L), 19.0; midshaft depth: WAM 09.3.284 (L), 7.0.
Phalanges: The paratype WAM 09.3.284 preserves five phalanges from the left foot (I.1, III.1, III.2, ungual?III.3, and IV.1) and os metatarsal I. The metatarsal (hallux) is about 20% longer than that of Ce. violaceus and proportionally similar. Phalanges 1 and 2 of digit III and phalanx 1 of digit IV are almost identical in length to those of Ce. violaceus and therefore much shorter relative to body size, but they are much wider and deeper and thus more robust. In the phalanges of digit III, the dorsal surface projects strongly proximally (in III.1 this projection articulates with the wide, deep groove in trochlea metatarsi III of the tarsometatarsus; in III.2 it articulates with a groove in the distal end of III.1) providing lateral stability in the toe. The ungual is markedly curved, and of similar length to that of Ce. violaceus and thus is shorter relative to body size (the unguals of the available Ce. violaceus specimen are covered with the keratin sheath, making comparisons difficult). Overall, the phalanges of Ce. maximus appear to have been proportionally shorter than in Ce. violaceus , with a smaller foot span relative to body size presumed to correlate with more terrestrial habits in the extinct species. The more robust phalanges of Ce. maximus are consistent with the toes supporting the weight of a larger, heavier animal.
Remarks
This species was the largest known cuculid from anywhere in the world, with measurements of the humerus and femur indicating that it outsized the previously known largest species, Ce. colossus , also from the Pleistocene of southern Australia. Centropus maximus had a tarsus length much greater than that of any extant member of the cuckoo family, including terrestrial species such as the Geococcyx roadrunners and Neomorphus ground-cuckoos of the Americas and the Carpococcyx ground-cuckoos of South-East Asia, and its long tarsometatarsus was matched by a very long and robust femur and tibiotarsus. The humeral morphology (curved, rela- tively slender shaft, much reduced caput humeri, and ventrally extended distal end) indicates that this species was poorly adapted to flight, and the very large and robust leg elements coupled with small feet suggest that this taxon was probably adapted to terrestrial locomotion, rather than to climbing.
Body mass and proportions
Body mass estimates: The eight selected regression equations of Campbell & Marcus (1992) produced a range of estimates for each of the three fossil species, with considerable variation within and amongst species ( Table 4): estimates for Ce. colossus ranged from 2.2–2.3 kg; Ce. bairdi from 1.1–2.1 kg; and Ce. maximus from 2.3–3.1 kg. To assess which equation produced the most reliable estimates for large cuculids, we applied the same calculations to specimens of extant species. The resulting estimates are graphed as a proportion of published mean body mass for each species ( Gilliard & LeCroy, 1967; Dunning, 2008; Erritzøe et al., 2012; Fig. 6 View Figure 6 ).
The equation based on nonpasserine tibiotarsus data consistently overestimated the body mass of extant cuculids whether they are highly volant (e.g. Scythrops , Eudynamys ) or minimally so (Ce. phasianinus, Ce. bengalensis, Ce. violaceus ) ( Fig. 6 View Figure 6 ). Thus we regard the maximum estimates for the two fossil species to which we could apply this equation as implausibly high (2.1 kg for Ce. bairdi ; 3.1 kg for Ce. maximus ). The long-legged bird tibiotarsus equation produced the best estimates for extant Ce. bengalensis and Ce. phasianinus (1.1 and 1.2 times known mean body mass, respectively). This equation also produced a highly accurate estimate for the individual S. novaehollandiae specimen we measured, which despite not being a long-legged taxon, had an estimated mass within 10 g of the actual weight recorded on its museum label. However, although it worked well for the smaller species, the long-legged bird tibiotarsus equation appeared to significantly overestimate the body mass of the largest extant species included in our comparison, Ce. violaceus , as did all other equations, signalling that they may also have produced overestimates for the very large extinct species.
However, the mean and range of body mass for Ce. violaceus are unknown. Its recorded weight (500 g) is based on only two individuals, the specimen reported in Gilliard & LeCroy (1967) (cited by Dunning, 2008 and Erritzøe et al., 2012) and the Australian Museum specimen used in our study (unpublished collection data). The skeleton of Ce. violaceus is far larger and more robust than that of Ce. phasianinus , for which individuals of> 600 g † Based on two individuals, both weighing 500 g.
are known ( Dunning, 2008; Erritzøe et al., 2012), and so a body mass of rather more than 500 g for at least some Ce. violaceus individuals seems intuitive. However, this species may be lighter than expected owing to reduced musculature of the pectoral girdle, which may also be true of the three extinct species (see Systematic palaeontology).
For smaller cuculids we regard the long-legged bird tibiotarsus equation as the one most suitable for estimating body mass. If this equation were also accurate for large species, then our best estimate for Ce. bairdi would be 1.6 kg, and for Ce. maximus 2.6 kg. This equation could not be applied to Ce. colossus , but three of the four femoral equations produced estimates of 2.2–2.3 kg for this species. Given the uncertainty about the suitability of any of these equations for very large coucals, the actual body masses of the extinct species may have been rather lower than those estimated. If 500 g is accepted for Ce. violaceus , then the long-legged bird tibiotarsus equation overestimated its mass by 2.46 times. Assuming that the same overestimate is true of Ce. bairdi and Ce. maximus , their revised estimates would be 664 g and 1 kg, respectively. Apply- ing a similar correction to Ce. colossus , based on a 2.17-times overestimation of the mass of Ce. violaceus using the all-species femur equation, its revised mass would also be around 1 kg.
Simpson log-ratio diagram: The postcranial body proportions of extant and extinct species of Centropus are shown in Figure 7 View Figure 7 , where they are graphed relative to the comparator species, the highly volant channel-billed cuckoo S. novaehollandiae . Differences amongst the species of Centropus are minor and their proportions are similar: all have shorter proximal (humerus) and distal (ulna and carpometacarpus) wing elements than S. novaehollandiae , a shallower keel on the sternum, elongate coracoids, wide femoral midshafts, and very elongate tarsometatarsi. The proportions of the extant taxa Ce. bengalensis and Ce. phasianinus appear especially similar to one another, as do the proportions of the extant Ce. violaceus and extinct Ce. maximus . The limited skeletal remains for Ce. bairdi and Ce. colossus indicate that they shared similar humeral proportions, but had dissimilar femoral proportions.
PHYLOGENETIC ANALYSIS
Executing Hughes’s (2000) original data matrix in PAUP* with the same settings and without any additional species recovered the original tree topology, so we were confident that the reconstructed data matrix was error-free. Since the publication of the original study, it has been established that the outgroup used by Hughes (2000) – Opisthocomus (Hoatzin) and the Musophagidae (turacos) – is not monophyletic (e.g. Jarvis et al., 2014). However, setting the outroot as paraphyletic or as a polytomy did not alter the topology of the original tree, indicating that the outgroup remains appropriate for polarizing the included characters.
Adding all the extant and fossil taxa into the original matrix produced a poorly resolved consensus of> 8000 shortest trees, but it placed Scythrops close to Eudynamys / Urodynamis as expected. Centropus phasianinus, Ce. violaceus , and the three Australian fossils were the only taxa besides Coua that did not join a clade within the polytomy (tree not shown). To determine the causes of poor tree resolution, we trialled adding different species combinations to the original data set. Using only extant cuculids without any fossils, the best resolution at the basal end of the tree was found when Ce. violaceus was included and Ce. phasianinus excluded, and thus Ce. phasianinus was temporarily excluded in order to further investigate the effects of the fossils upon tree topology.
Simultaneously adding all three fossil species produced a polytomy, as did adding Ce. bairdi and Ce. colossus individually. However, adding Ce. maximus alone produced a resolved consensus of only four shortest trees, in all of which Ce. violaceus and Ce. maximus formed a clade between Carpococcyx and Coua . It is likely that the poor resolution seen with the inclusion of Ce. bairdi or Ce. colossus is because of the limited number of skeletal elements that we have for each (~5% of scored characters for Ce. colossus ; 18% for Ce. bairdi ). The more complete remains of Ce. maximus , by contrast, allowed scoring of 40% of all characters, and 57% of postcranial characters. We therefore excluded Ce. bairdi and Ce. colossus from further analysis in order to examine the affinities of Ce. maximus .
With Ce. bairdi and Ce. colossus thus excluded and Ce. phasianinus reincluded, the resultant consensus of 16 shortest trees ( Fig. 8 View Figure 8 ) was less well resolved than when Ce. phasianinus was excluded. It nevertheless retained similar topology to the tree of Hughes (2000), with the bootstrap analysis confirming monophyly of the Cuculidae with 100% support. Scythrops novaehollandiae , which was not included in the original phylogenetic study, joined the clade that Hughes designated as the ‘Cuculinae’, albeit with dramatically lowered bootstrap support for that clade compared with the earlier study (52 vs. 98%). The ‘Phaenicophaeinae’ of Hughes was slightly less well supported (52 vs. 54%) with Rhinortha no longer falling within the clade. The ‘Crotophaginae’ retained 100% support, as did the ‘Neomorphinae’. The ‘Centropodinae’, within which Hughes included Centropus and Coua , collapsed into a polytomy, but within this there was strong (90%) support for a clade formed by the extant Ce. violaceus from Melanesia and extinct Ce. maximus from the Nullarbor Plain. Centropus phasianinus did not join this clade, and Carpococcyx continued to form its own clade separate from Coua and Centropus with 100% bootstrap support.
Variations in the arrangement of the branches for Carpococcyx , Coua, Ce. phasianinus, Ce. violaceus , and Ce. maximus within the 16 shortest trees from which the consensus tree is derived are summarized in Figure 9 View Figure 9 , with the branch orders shown at A, B, C, and D having been recovered four times each. The position of Carpococcyx remained stable in all trees, whereas the remaining four ingroup taxa were arranged four different ways. Coua and Ce. phasianinus were each other’s nearest neighbours in all trees, forming a clade in a quarter of trees ( Fig. 9C View Figure 9 ). The extinct Ce. maximus and extant Ce. violaceus formed a clade in all trees, although the position of this clade varied in relation to Coua and Ce. phasianinus : in 50% of trees, Ce. phasianinus was the nearest neighbour to the Ce. violaceus / Ce. maximus clade ( Fig. 9A, D View Figure 9 ); and in 75% of trees Ce. phasianinus was equally close or closer to the Ce. violaceus / Ce. maximus clade than was Coua ( Fig. 9A, C, D View Figure 9 ).
In the bootstrap consensus tree ( Fig. 8 View Figure 8 ), the Ce. violaceus / Ce. maximus clade was supported by 13 apomorphies, of which only three were unambiguous: Character 69 [tarsometatarsus, position of tuberositas m. tibialis cranialis: distinct gap between the tuberositas and foramina vascularia proximalia, consistency index (CI) = 0.250, 0 ==> 1]; Character 96 (sternum, form and fusion of rostrum sterni: spina interna and spina externa absent, CI = 0.833, 0 ==> 5); and Character 97 (sternum, size of spina externa: absent, CI = 1.000, 0 ==> 2).




| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.